|
|
|
The main aim of this book is to describe the Swedish and Finnish reintroduction projects in some detail. Background information is given in the papers on the population history and other conservation measures. The articles on probable future scenarios intend to give some tools to the reader who wishes to estimate the effects of future action alternative
Martti Soikkeli
When the need to rescue the Lesser White-fronted Goose became apparent in the 1960s, nobody really had an idea of what the main threats to the geese were and what should be done.
Fifteen years later, Lambart von Essen was convinced that the problem was high hunting pressure on the south-eastern migration routes characteristic to the species, and it was both necessary and possible to shift the migration routes to safe areas in Western Europe. He was indeed able to create a new migration route by putting LWfG eggs into nests of Barnacle Geese. Later on the Lesser White-fronted goslings would migrate to the Netherlands in company of their foster parents. They were imprinted on the new route and survived - for generations. Lambart's method really was a success, and today his geese are naturally multiplying in Sweden by almost 10 per cent annually. Last winter, more than 90 of them were wintering in the Netherlands. On the other hand, the original population died out both in Finland and Sweden, and the prognosis is hopeless for the below 10 observed and below 25 assumed pairs still breeding in Norway.
In 1998, traces of hybridisation with (greater) White-fronted Geese (Anser albifrons) were detected in the genome of some of the captive birds. Releases were interrupted and gene test samples were taken from essentially all the 250 captive birds in Sweden and Finland, more than half of the world's captive Lesser White-fronts. Active conservation in nature was reduced to supporting the remaining Norwegian population by investigating conditions along their Eastern migration routes - and hoping to improve them some time in the future. Careful observations did indeed lead to some new insight like the discovery of a previously unknown branch of the autumn migration flyway going directly east over the White Sea. In fact this seems to be the only remaining route, since fall observations in Finland have ended completely a decade ago.
Two groups of conservationists have emerged. Some would like to fix criteria for the genetic suitability of the goslings and then continue von Essen's project. The other party seems to believe in a more or less natural recovery of the Scandinavian population. In my opinion, these two must find a way to compromise and cooperation.
As an old LWfG-fan I would regret extinction of the LWfG in Europe. I do not consider the genetic findings an impenetrable barrier against the reintroduction programs. The observed differences - scientifically interesting as they may be - are too small to have any effect on the appearance, behaviour or survival rate of these birds. It is also well known that there is a steady gene flow between closely related waterfowl species in nature, and this is compensated by other mechanisms keeping the species apart. Also, the "Norwegian" birds spend most of the year on migration and at their wintering sites, where any female may pick up some arbitrary male with "Siberian" genes to become father of our "purely Scandinavian" geese, so there is another gene flow between geographically distinct LWfG - possibly except the western and eastern sub-populations as defined in recent comprehensive studies (Vladimir Morozov).
Soon after the interruption in the Swedish project, somebody presented the extreme opinion that all geese originating from Lambart von Essen's reintroduction program should be shot. Fortunately, this will not be done, and most captive birds are kept alive as well. Lambart von Essen himself once spoke the wise words: "These birds look like Lesser White-fronted Geese. They behave like Lesser White-fronted Geese, and they are perfectly adapted to arctic Sweden. Why should they not be Lesser White-fronted Geese!"
Johan Mooij
Anser erythropus nests in tundra with none or low tree density at lakes or near rivers. Nests are built in grassland, between dwarf shrubs or in hollows in wetlands. Anser erythropus is an exclusively palearctic breeding species. Nestlings hatch after a breeding period of 25-29 days in the first third of July. Average number of eggs per breeding pair is about five. Except for newly hatched goslings, A. erythropus are strictly herbivorous and feed on grasses and herbs on riversides, lakeshores and swamps. Main predators in Fennoscandia are Vulpes vulpes, Alopex lagopus, Aquila chrysaetos, and Haliaaetos albicilla.
Edited by Lauri Kahanpää
Adapted from [Nor] and [LIFE], [P]:
The original range of the Fennoscandian population of the Lesser White-fronted Goose (A. erythropus) covered the arctic parts of all Fennoscandia.
Migrating geese arrive at breeding grounds about end of May and leave about end of August. Parts of the Fennoscandian population migrate via the Baltic Region and Hungary to the Balkans. A second route leads via western parts of Russia to the Black sea, while a third route goes east through Kazakhstan to wintering areas around the Caspian Sea. Few are observed in Germany and The Netherlands.
In 1920-30, Erkki Merikallio mentions tenthousands of A. erythropus on autumn staging sites at the Bothnic gulf and an original Finnish population of 6,000-10,000 individuals.
The Fennoscandian population was still estimated at about 10,000 birds in the 1950:s but a dramatic reduction had already set in. In the seventies, no more than 100 pairs seem to have been breeding, and by the middle of the 1990:s the population was down at 25-30 breeding pairs (26 in 1997).
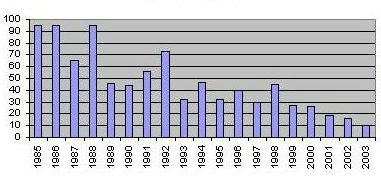
Spring observations in Oulu area, Finland Today, no regular autumn migration is observed at the Bothnic Gulf any more. In spring, only the traditional staging sites near Oulu are in use. In twenty years the number of A. erythropus observed there has fallen from 100 to 10 - an average annual decrease of 11 per cent.
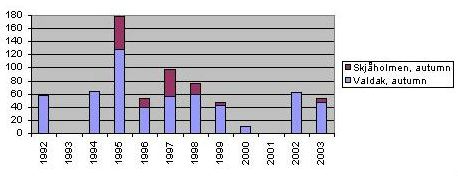
Autumn observations in Norway, 2 sites There are two known autumn staging sites in Norway. Also direct counts in the breeding area have been carried out (in Finnmarken, Norway. Earlier breeding areas in Finland, Sweden and - most recently - Nordland in Norway are empty.). The overall picture shows relatively large fluctuations in the breeding success overlapped with an overall down trend of 5 per cent annually.
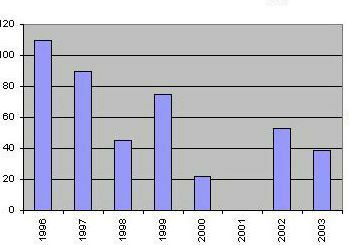
Early spring observations in Hortobagy, Hungary Wintering Scandinavian A. erythropus are difficult to count, since they mix with Russian birds. But there is an early spring staging area (Hortobagy) in Hungary. Available numbers from there give an annual decrease of 11 per cent.
Johan Mooij
The reproduction rate of Anser erythropus where it is still breeding, is at normal level compared to other arctic geese: an average of 40 per cent young birds in autumn is observed. But mortality is extremely high, in particular for young geese (annual 70-80%). Natural predation seems to play a minor role except in years of an extremely low offer of small mammals. Instead, hunting seems to be the most important factor, as 50% of the young geese and 10% of adults marked in Norway were reported to be shot. Anser erythropus resembles Anser albifrons and often occurs in mixed groups with this common and heavily hunted arctic goose species. Therefore, a significant number of A. erythropus are shot in spite of world wide protection.

A. albifrons, shot in Olonets, Russia. (Foto Erkki Kellomäki)
Due to the immense negative impact of hunting, the Fennoscandian population of Anser erythropus has decreased to a population size small enough to risk a total loss due to e.g. diseases or a series of subsequent years with low reproduction rates. The high probability to find a relative as mating companion leads to inbreeding risks by further loss of genetic diversity. Appropriate ecosystems along the traditional migratory route as well as in the wintering area in the Caspian region have also been subject to major changes. Wetlands are dried out and meadows are taken into human use.
Vladimir Morozov (Abbreviated from [Caz 8])
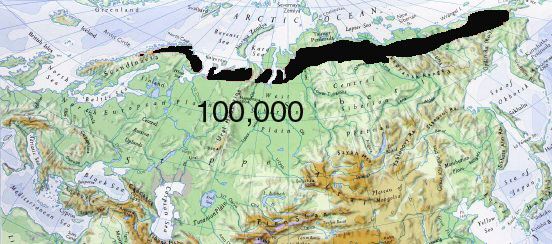
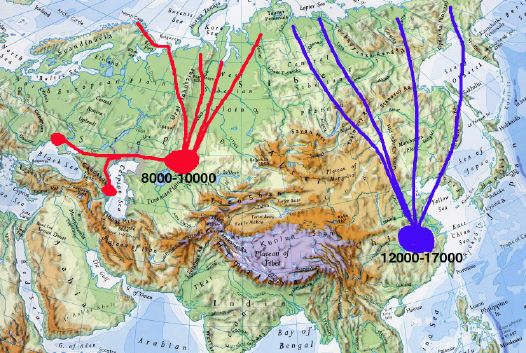
Fifty years ago, the breeding range of the Lesser White-fronted Goose was a connected belt reaching from the Atlantic across arctic Eurasia to the Pacific. The species is estimated to have numbered some 50,000 pairs. (No Anser erythropus breed in the Nearctic.)
Anser erythropus range ca 1920 Since the last review of the world population of the Lesser White-fronted Goose [Morozov 1995], a great amount of data was accumulated. Today, the breeding distribution and numbers can be estimated separately for each geographical region.
Since the first estimates of the size of the world's population in the 1950:s, a drastic decrease in numbers was recorded from more than 100,000 individuals in the 1950:s to less than 25,000 birds in the 1990:s.
In particular, in the Western Palearctic flocks of c. 50,000 A. erythropus were regularly recorded during the 1930:s, and during the 1950:s the Western Palearctic population was still estimated at more than 50,000 birds. In the 1990:s numbers recorded in this region during midwinter counts never exceeded 10,000.
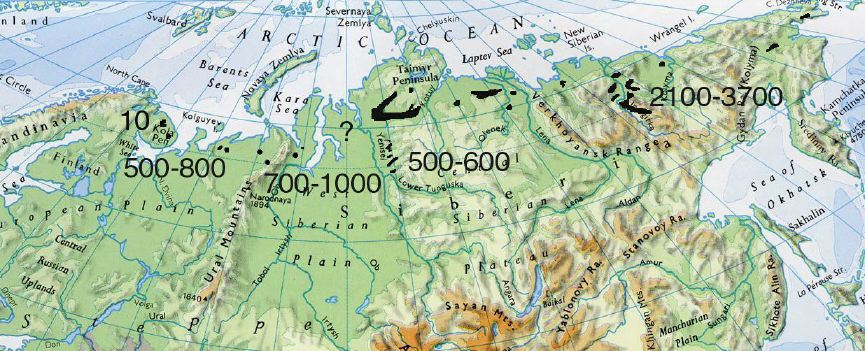
Anser erythropus known range today We attempted to determine the numbers of every territorial group inhabiting particular geographical region within the breeding range of the species. We did not succeed completely, because the breeding numbers could not be evaluated for all known regions. The spring numbers of the species was estimated:
- European Russian tundras: 500-800 adult birds
- Yamal Peninsula: 700-1,000
- Taimyr: 2,000-3,000
- Putorana Plateau: 150
- Abiskyaya Lowland in Yakutia: 2,100-3,700
The total numbers approximates 4300-5900 individuals at the beginning of the breeding season: in autumn this would yield about 6,000-8,500 individuals. These numbers are lower estimates. We were not able to evaluate the present numbers of the species in some regions undoubtedly inhabited by the Lesser White-fronted Goose. (Gydan, Taimyr, many areas of Yakutia and Chukotka). On the other hand, data of goose censuses on the wintering grounds and migration routes is available.
A. erythropus migration Current estimates at stopover sites in Kazakhstan arrive at present numbers of the species in West Siberia from 8,000 to 11,000 individuals. If most of the European birds arrive to Kazakhstan for ret and fattening, the total numbers of the western half of the overall population can be estimated at 9,000 to 11,000 individuals in autumn.
We have neck-ringed 16 adult and 4 juvenile LWfG in the Polar Urals in 1999-2002. The only resightings were 2 juvenils shot in the same autumn (2000) in Maynach Valley, and a third one killed in Tyumen Oblast (West Siberia) same season. So, the question about the exact migration route of the geese of the Polar Ural region remains unsolved. However, it was confirmed that the Maynach river Valley remains one of the key stop-over sites of the LWfG during migration. Unfortunately, we do not know, where the wintering grounds of this population of LWfG are located. We are only able to guess that they can be found somewhere in South-Eastern Europe, because the ringed birds were shot near the Azov Sea belonging to the Mediterranean basin.
The only east Asian flyway known in some detail is the easternmost one on the map. The eastern population is known to winter in China, mainly at Dongting Lake in the lower Yangtse area. Here 16,600 individuals were found in January 2003 [B]. This would yield a global autumn population of 28,000 individuals.
About 80 per cent of the original population have disappeared since 1920. During the very last decade, no abrupt change was observed, only a continuous decline in numbers, in particular of the Fennoscandian population. The losses of marked birds during migration vividly demonstrate high mortality in this population after the breeding season. Increased mortality, mainly due to hunting pressure along migration routes is considered virtually the only cause of the number decline. However, also the Siberian populations suffer from both legal and illegal hunting. Breeding results have been monitored. Normal numbers are 3,5 to 4,5 juv / successful breeding, depending on the year.
New data do not confirm the existence of a number of distinct geographical populations. On the contrary, the results of satellite tracking of birds marked in Fennoscandia and ringing in the Polar Urals favour the concept of the existence of one single population inhabiting the territories from the north of Scandinavia to the east of Taimyr. It is suggested, that a second population inhabits the territory from eastern Taimyr to Chukotka, since a vast majority of these birds winter on a comparatively small area in China.
Lauri Kahanpää
The Lesser White-fronted Goose is extinct in my home country, Finland. Reintroductions are one way out of the hunting pressure problem. But are they really necessary? What are their implications? To estimate the future, we have to know the facts. So let us list some facts. Details are found in the papers in this Bulletin:
- The White-fronted Goose is globallly threatened, almost extinct in Scandinavia.
- The reintroduced Swedish population is growing, in the average by at least 5 per cent.
- A Finnish program failed, since it did not use foster parents.
- In chromosome DNA, differences between captive and wild Lesser White-fronts are negligible. In mitochondrial DNA, captive and wild geese have some differences, the most serious being indications of hybridization with White-fronted Geese (Anser albifrons).
- The new Swedish population is genetically identical to the captive one.
- The remainders of the Norwegian population show an average annual decline of at least 5 per cent.
What happens if current trends continue? To foresee the future of the Norwegian and Swedish birds, we can simply believe in exponential growth of non-saturated populations. The author has built some more elaborate mathematical models taking into account variations in breeding success in relation to the bird's age and to yearly weather conditions, and compared the results with observations. It turned out that the effects of these factors on the overall predictions were negligible. Variations in mortality during migration could have a negative effect on the population estimates, but these were not included in models, since there exists no sufficient data to work on.
But the future of the Lesser White-fronted Goose in Finland does not only depend on these trends, since the reintroduction programs can be resumed. Below, their effect on the number and genetic composition of the future birds is calculated.
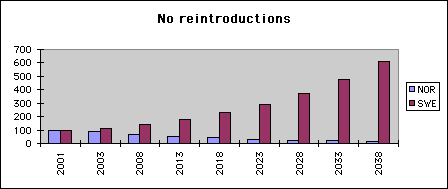
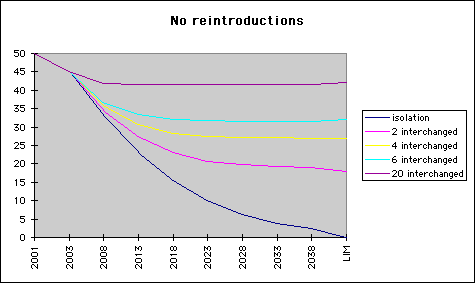
Diag.1. Growth and decay of 5% Let us display the effect of an annual 5 per cent change on the population over a time span of almost 40 years. In the diagram, we start with 100 birds - a reasonable approximation of the real starting situation - and follow the size of a population assuming a 5 per cent decrease "NOR" or a corresponding growth "SWE". Saturation will not be reached in the time span.
Let us now think, the populations are genetically different, and see what will happen if nothing is done. Diagram 1 gives the impression, the NOR type genes will vanish. Fortunately this need not necessarily be the case.
Diag.2. Average percentage of NOR genes in Scandianvian population
The curves in diagram 2 give the ratio of numbers of NOR to all birds. (LIM stands for the theoretical result in a very long time perspective.) The lowest curve displays the ratio of NOR to all birds in diag 1, and converges to zero, as expected. In the case of isolated populations this is the ratio of NOR genes in the total Scandianvian stock. The other curves describe the result of gene flow between both populations. Flow is modeled by interchanging one, two, three or 10 pairs of adult birds annually, and - for simplicity - assuming homogeneity and instant diffusion of genes in each population. The effect will - of course - be a flow of genes from NOR to SWE, where they will be preserved. An opposite flow will dilute the NOR genes in the NOR population, and will - by time - lead to equilibrium at a level visible in the diagram. By the time of essential extinction of population NOR, the gene flow will of course halt, but we can read from diagram 2 that equilibrium is essentially reached already before - on a level solely depending on the intensity of the gene flow.
An open question is of course, how much gene flow there is in reality and what could be done to increase it in order to preserve much of the NOR type.
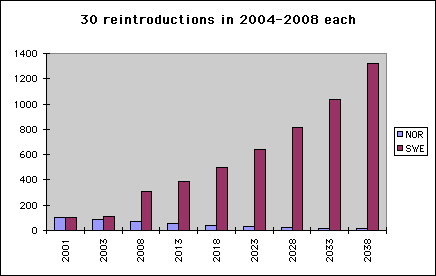
Diag.3. Growth and decay of 5% + restocking Diagrams 3 and 4 show the same things as diagrams 1 and 2 with one difference: This time we assume that 30 type SWE birds are annually added to population SWE during five consecutive years. A period of extra growth in SWE population numbers is visible in diagram 1., and the final population is doubled. NOR numbers are not changed.
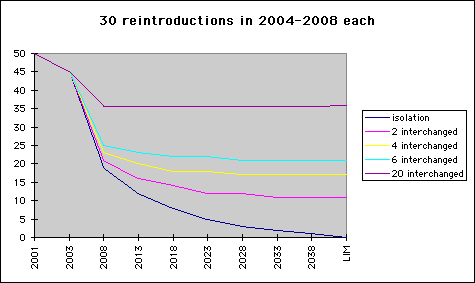
Diag.4. Percentage of NOR genes in Scandianvian population with restocking
Of course, the immediate effect of reintroductions by SWE type birds is an increase of SWE type genes. Another effect is that equilibrium is reached earlier.
The effects of the assumed restocking are:
- The numbers of SWE type birds will double quickly. This will reduce the risk of losing not only NOR but also SWE.
- SWE will sooner reach a level of 200 pairs (600 birds) - a size large enough for ending restocking.
- Carrying out some -or all - of the restocking with NOR birds would increse the ratio of NOR genes in the final (LIM) stock.
- The same effect can be reached by incresing gene flow between both populations.
- Let us also note in passing:
- The natural gene flow can be increased by carrying out the restockings close or inside the breeding area of NOR.
- Nonhomogeneity of each population is unknown.
- Delaying reintroduction to 2005-2009, say, will reduce all its effects, including the preservation of NOR genes. (No diagram drawn).
- The possible effects of slow diffusion are not estimated here.
- If some of the genes in NOR or SWE are favourable, they will be enriched by the Darwin process, unless they appear only in NOR and there is no sufficient gene flow from NOR to SWE.
- The differences between NOR and SWE are unknown - they may be negligible.
The Lesser White-fronted Goose is a globally threatened species that is classified as Vulnerable (VU). It is formally protected by law in essentially all relevant countries.
Descriptions of detailed actions and results is given in the articles in [LIFE]. Here we just list some further points of interest in the context of reintroductions and other activities of the Friends of the Lesser White-fronted Goose.
- In the second half of the 1990:s, the EU Commission approved an International Action Plan for the conservation of A. erythropus. The Action Plan plan has not been updated since then.
- To keep track of and coordinate research and conservation measures, Wetlands International's Goose Specialist Group has a special organization.
- Since 1997, Häme Regional Environment Centre (Finland) is doing remarkable co-operation with authorities in Russia and Caspian Sea countries in order to increase awareness of national authorities on vulnerable situation of this species and reduce their mortality.
(Åke Andersson and Torsten Larsson)
In the late 1970:s the population of the Lesser White-fronted Goose in the mountain area of Swedish Lapland had been reduced to very few pairs. In 1981 the Swedish Association for Hunting and Wildlife Management with support from World Wildlife Fund, Sweden, started to release Lesser White-fronted Geese in one site within the former breeding area in Lapland.
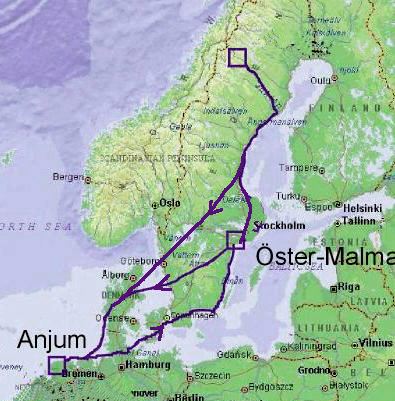
A captive stock of LWfG is kept at Öster-Malma Wildlife Management School in central Sweden. Young are reared at Öster-Malma and released in Swedish Lapland with Barnacle Geese (Branta leucopsis) as foster parents. The goslings are imprinted on the release area and are guided by their foster parents to winter quarters in The Netherlands. In spring the immature LWfG geese accompany the Barnacle Goose back to their breeding area in central Sweden but continue alone to the area in Lapland, where they were released.
On their spring migration from the Netherlands, the Lesser White-fronted Geese use a number of stopover sites along the coast, but during the autumn migration only two sites in central Sweden are regularly used. From there most geese seem to fly directly to the winter quarters in the Netherlands. The original population of the Lesser White-fronted Goose migrates to the Caspian/Black Sea regions, where significant hunting pressure is an alarming problem. By changing the migration route fir the Swedish population from southeast to southwest, the geese are offered a much safer staging and wintering area. In 2003, the total Swedish population was estimated to about 100 individuals.
Up until 1999, a total of 301 goslings and 47 one-two year old geese were released in the mountain area of Swedish Lapland. (Foto Erkki Kellomäki) At least 53 breedings have been confirmed. More than 47 breedings have resulted in fledged young recorded at staging areas along the coast of Gulf of Bothnia. During the last four years 25 broods with a total of 70 immatures have been produced, 2002 being the best year producing 8 broods and 21 young geese
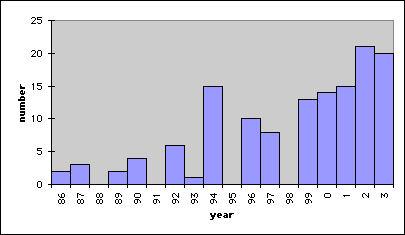
Number of fledglings recorded from introduced geese or their decendants. In 1995 no breeding took place because of cold weather and in 1998 the ice broke up very late, causing high nest predation.The average growth rate of 5 per cent is maintained since 1999 also.
The genetic investigations on Lesser White-fronted Geese have confirmed that some of the birds in the Öster-Malma stock were contaminated with genes from White-fronted Goose (Anser albifrons). Pending the outcome of the genetic studies, the release of birds to Lapland has been temporarily stopped. Contaminated birds have been removed and the remaining stock of Lesser White-fronted Geese at Öster-Malma is being subject to pedigree analysis and estimation of relatedness to evaluate possible future use for reintroductions.
Pentti Alho, Lauri Kahanpää and Erkki Kellomäki
The Swedish reintroduction project has had some success while a simultaneous Finnish project has failed. Why? Learning from experience is important for many reasons - one of them the economical reason: The total costs for one A. erythropus gosling rise as high as 1000 EUR. Now - 20 years after the start of the projects - it is time to sum up our cumulative experience:
During two decades, about 350 LWfG were set free in Sweden's arctic mountains. Today these birds breed and are in the process of forming a self-sustained population in spite of the temporary release interruption.
Finland started a similar project eight years later. All in all 150 goslings were released in Finnish Lapland but not a single breeding of these birds has been recorded. The last known breeding was an original pair in 1995.
In Sweden, fertilized LWfG eggs were taken away from their natural parents and moved into nets of free-living Barnacle Geese (Branta leucopsis). Later, the freshly hatched goslings were caught together with their foster parents and taken to the nearby bird farm. The goslings were imprinted on the Barnacle Geese.
In Finland, captive goslings were imprinted on their natural parents without human interference. There were no other major differences between origin and conditions for captive birds in Sweden and Finland.
In Sweden, the Barnacle Geese parents, together with their LWfG goslings, were transported to the Arctic by car and helicopter. This was done, when the goslings were 6 to 7 weeks old, just learning to fly and to escape predators.
In Finland, the LWfG families - in many cases only the female with the goslings - were transported to Lapland by car and plane when the goslings were only 1-3 weeks of age. In 1997, a released family was carefully observed in their first fortnight in freedom. During this time, a red fox killed the female and all the goslings.
Fall migration of the Swedish geese was easy to observe, since the probable staging places were known in advance. The Barnacle Goose parents favour riverside meadows close to human settlement in the Bothnic Gulf region. During their migration round trip, the survival rate of the LWfG goslings was an impressive 80 per cent.
In Finland, the released birds were observed for some time near the release site. Later they disappeared until some were seen in winter scattered in various countries. Possibly these birds have joined flocks of migrating Anser fabalis. Of the Finnish project geese only one individual has been observed in spring in Inari, Finnish Lapland. No breedings are known
Pentti Alho
Breeding Lesser White-fronted Geese in Finland was begun in 1986 by importing two pairs from Sweden to a farm on the island Hailuoto (Karlsö) in the Bothnic Gulf. Already the same year, the first five goslings hatched - the next year more. Releases were carried out since in 1989 and a second goose station was constructed in Hämeenkoski near Lahti in south Finland. Unfortunately, bird tbc struck the Hailuoto farm, which was closed in spite of a successful disinfection program in 1998.
The station in Hämeenkoski received their first birds from Hailuoto but was spared losses by infectuous disease. Today our farm is the largest Anser erythropus breeding station in the world housing about 110 gene tested Anser erythropus - about one third of the world's captive population. Separate breeding compartments for each pairs have been in use since 1994. Ongoing construction work has fundamentally improved all farm facilities during our operation years.
In 1998, while searching for differences in mitochondrial DNA in Lesser White-fronted Geese from different localities, the geneticists Jaakko Lumme and Minna Ruokonen studied birds from the Hailuoto stock. They discovered some haplotypes close to haplotypes of the White-fronted Goose (Anser albifrons), an observation which suggested some hybridization between the two species in the past. Follow-up studies led to similar observations in Sweden.
Mitochondria are inherited strictly maternally. Since mitochondria do not carry genes, geese with different mitochondrial DNA look and behave alike, and without tests it is impossible to find out which individuals bear which haplotypes. To clarify the situation, the chromosomes themselves must be compared. Finnish samples were analyzed at the Servetsov Institute for Ecology and Evolution - a branch of the Russian Academy of Science in Moscow. Decisive signs of hybridization were not found in the chromosome DNA. The need to use even more sensitive tests is under disussion. On the other hand, it is known - and new information at Wetlands International's Odessa meeting confirmed - that Lesser and Greater White-fronted Geese (A. albifrons) do hybridize in Nature, and that their offspring are fertile.
According to IUCN guidelines, best available stock should be used for reintroductions, preferring remnants of a local population, if there is any. Enriching or replacing our captive stock with wild geese must be considered seriously. However, a supplement of pairs of wild Norwegian or Russian Lesser White-fronted Geese is difficult and expensive to acquire. Single birds taken from nature rarely breed in captivity. For that reason, we have to take good care of the genetic diversity in our captive stock.
Lesser White-Fronted Geese breed in separate pens with one pair in each. (No foto during breeding!)

At Hämeenkoski, two independent flocks can be housed all year round in separate enclosures, each comprising of a lawn and a winter hall.
Aktion Zwerggans
(Foto Aktion Zwerggans) In 1999 a French team led by Paola and Christian Moullec bred a group of A. erythropus which were imprinted on a microlight airplane. The young geese were transported to Sweden by car. In autumn they were guided by the microlight plane to Bislicher Insel, a well known and protected wintering area of arctic geese in the lower Rhine area in Germany. There they were released by draw-back of the plane. With a return-rate of 40 - 50 % of the released A. erythropus in middle-Sweden, the test project proved to be rather successful. This return-rate is comparably high as in the Swedish reintroduction project.
As a side effect, the test flight of 1999 triggered an incredible engagement and public awareness for the conservation problems of wild geese.
In 2001 Aktion Zwerggans (www.zwerggans.de) was founded in Germany. Aktion Zwerggans has improved the microlight method according to latest scientific and technical knowledge (LIFE Starter project LIFE2002NA T/ST/D/000005) and is prepared to implement this method within Fenno-scandia.
Lauri Kahanpää
In March 2004, Wetlands International's Goose Specialist Group had their annual meeting in Odessa, Ukraine. Since the the next meeting will be in Hungary in autumn 2005 - two breeding seasons from now - another meeting, dedicated solely to the Lesser White-fronted Goose, will be arranged in spring 2005. A perfect place for this is Finland.
The Friends of the Lesser White-fronted Goose wish all conference members welcome in Helsinki.
To the remaining wild A. erythropus population in Scandinavia, the main threat is excess hunting along their migratory routes and in wintering areas. An effective control of hunting along the migration routes scattered all over Russia is of course important, but very difficult to achieve in a foreseeable future. This was confirmed by the representatives of Russian hunter's organizations at the 2003 meeting of the Goose, Swan, and Duck Study Group of Northern Eurasia (RGG) in Olonets, Russia. Therefore, most emphasis should be laid on improving condition in the Caspian Sea region, a very important staging and wintering area of A. erythropus.
The Friends of the Lesser White-fronted Goose will do all we can to promote research and the set up of action plans in Kazakhstan, Uzbekistan, Turkmenistan, Iran and Azerbaijian. Fortunately, a large percentage of our members are fluent in Russian.
The reintroduced Swedish population may face an unexpected threat in the near future as the Netherlands are reopening goose hunting. No localities where Anser erythropus are observed should be opened for hunting.
Changing the migration route and wintering area of the reintroduced A. erythropus will essentially stop loss by hunting of these birds and their future offspring. Two methos should be considered:
Observations of all Lesser White-fronted Geese, both natural and re-introduced, are necessary for correct protection measures. This includes - of course - continued intensive monitoring in the breeding areas both in Norway and in Sweden. Also further research concerning the geese breeding in Siberia as well as further attempts to clarify their migration patterns should be supported.
In the reintroduction projects (old and future), the geese are marked with colour rings. Regular counts are carried out in Sweden and along the routes to the Netherlands. When reitroductions are resumed, it will be necessary to follow the moves the new birds. As a wide range of unoccupied potential breeding areas exist in Northern Fennoscandia, satellite telemetry provides the only possibility to do this effectively, in particular in spring, when the birds return.


SPOT3 is an almost prototype transmitter, specially designed to operate under difficult ocean conditions. Half of the global A. erythropus population migrates after the breeding season from the Siberian tundra through the Caspian Sea countries and winters there. Satellite data has already revealed that also parts of the vanishing Scandinavian population migrate to and through Caspian Sea countries, in particular Kazakhstan, where they are hunted. Satellite telemetry can help national scientists and authorities monitor the development of the current population. It can also be utilized to raise awareness among hunters and local people for the need to protect these geese. Western organzations - including the Friends of Lesser White-fronted Geese - must help to finance the purchase of telemetry equipment.
Satellite tracking will first be used to monitor the round trip migration routes of our Barnacle geese (Branta leucopsis) parent birds .The Friends have purchased two ARGOS satellite transmitters, type Wildlife Computers SPOT3. These have extended life, since they can be programmed to low intensity off migration time.
Couldn't we just remove the threats?
- No changes in hunting along migration routes in Russia can be expected in an overseeable fututre. This was clearly stated by the Russian hunter's organizations at the 2003 meeting of the Goose, Swan, and Duck Study Group of Nortern Eurasia (RGG) in Olonets, Russia.
- Protecting the species is impossible, since it looks like the Greater White-fronted Goose. Protecting wintering places is impossible for the same reason - also yearly variations in humodity force the geese to vary winering lakes, so there seeemingly exists no particular LWfG-place to protect.
- The Lesser White-fronted Goose would still return spontaneously to Scandinavia, if hunting ceased in the east - but that will not happen.
Why are the Greater White-fronts doing so much better?
- Most of them winter in Western Europe. But A.albifrons are heavily hunted along their migration routes, too - in fact they are the number one hunted Goose species in Eurasia, and recent data (Odessa 2004) points at a change in their positive population trend.
Why the hurry?
- It can be inferred from the estimates in the "scenarios" paper bove, that the optimal time to start restocking passed already. Prolonging the interruption will lead to a higher risk of losing one or both of the Scandinavian populations. Both are small, and (in contrast to birds of prey) geese are flock birds, having surplus mortality when their number goes down. (Similar effect as with the Passenger Pidgeon!). Also, the limit amount of recoverable "wild" genes decreases year by year.
Why don't we restock with original birds?
- We do not have any. No birds are available in Norway any more. No birds are yet available in Norway.
- Also, increased genetical varianbility is beneficial for the birds. Variability is the material for evolution.
- Starting breeding from scratch would take too long time to produce the goslings needed. (See previous answer)
- In contrast to other recent reintroductions, here the birds are gene tested for suitability.
- Ultimately, we must rescue the species.
Is it ethical to manipulate the natural behaviour of free-living birds?
- Yes. Changing migration routes is not unnatural - in fact it happens naturally all the time. Norwegian Horned Larks also migrate to the South-east - and are going extinct. Except for a small subpopulation going to the South-West. Also, the Red-brested Goose have spontaneously - by natural selection, that is - changed their wintering areas as well as their migration routes.
Why care for Europe? Are there not enough LWfG in Siberia?
- We care for both, but every country should primarily take care of their own biodiversity. (To tell the truth:: we just love to see them here!)
Will we follow the IUCN recommendations?
- Yes
What will happen with the LWfG in a) Norway b) Svaipa?
- Read the "Scenarios" paper.
Do we compete with other projects for the money?
- No. The Friends of the Lesser White-fronted Geese always prefer cooperation to rivalry.
How can You support the Projects
- Depends on Your skills and home area. Volunteers are welcome - material support as well. Breeding A. erythropus is a costly hobby, and satellite telemetry equipment is very expensive indeed!
Prepared by the SSC Re-introduction Specialist Group *
Approved by the 41st Meeting of the IUCN Council, Gland Switzerland, May 1995
These policy guidelines have been drafted by the Re-introduction Specialist Group of the IUCN's Species Survival Commission (1), in response to the increasing occurrence of reintroduction projects worldwide, and consequently, to the growing need for specific policy guidelines to help ensure that the reintroductions achieve their intended conservation benefit, and do not cause advers side-effects of greater impact. Although IUCN developed a Position Statement on the Translocation of Living Organisms in 1987, more detailed guidelines were felt to be essential in providing more comprehensive coverage of the various factors involved in reintroduction exercises.
These guidelines are intended to act as a guide for procedures useful to reintroduction programmes and do not represent an inflexible code of conduct. Many of the points are more relevant to reintroductions using captive-bred individuals than to translocations of wild species. Others are especially relevant to globally endangered species with limited numbers of founders. Each reintroduction proposal should be rigorously reviewed on its individual merits. It should be noted that reintroduction is always a very lengthy, complex and expensive process.
Re-introductions or translocations of species for short-term, sporting or commercial purposes - where there is no intention to establish a viable population - are a different issue and beyond the scope of these guidelines. These include fishing and hunting activities.
This document has been written to encompass the full range of plant and animal taxa and is therefore general. It will be regularly revised. Handbooks for reintroducing individual groups of animals and plants will be developed in future.
CONTEXT
The increasing number of re-introductions and translocations led to the establishment of the IUCN/SSCSpecies Survival Commission's Re-introduction Specialist Group. A priority of the Group has been toupdate IUCN's 1987 Position Statement on the Translocation of Living Organisms, in consultation withIUCN's other commissions.
It is important that the Guidelines are implemented in the context of IUCN's broader policies pertainingto biodiversity conservation and sustainable management of natural resources. The philosophy forenvironmental conservation and management of IUCN and other conservation bodies is stated in keydocuments such as "Caring for the Earth" and "Global Biodiversity Strategy" which cover the broadthemes of the need for approaches with community involvement and participation in sustainablenatural resource conservation, an overall enhanced quality of human life and the need to conserveand, where necessary, restore ecosystems. With regards to the latter, the re-introduction of aspecies is one specific instance of restoration where, in general, only this species is missing. Fullrestoration of an array of plant and animal species has rarely been tried to date.
Restoration of single species of plants and animals is becoming more frequent around the world.Some succeed, many fail. As this form of ecological management is increasingly common, it is apriority for the Species Survival Commission's Re-introduction Specialist Group to develop guidelinesso that re-introductions are both justifiable and likely to succeed, and that the conservation world canlearn from each initiative, whether successful or not. It is hoped that these Guidelines, based onextensive review of case - histories and wide consultation across a range of disciplines willintroduce more rigour into the concepts, design, feasibility and implementation of re-introductionsdespite the wide diversity of species and conditions involved.
Thus the priority has been to develop guidelines that are of direct, practical assistance to thoseplanning, approving or carrying out re-introductions. The primary audience of these guidelines is,therefore, the practitioners (usually managers or scientists), rather than decision makers ingovernments. Guidelines directed towards the latter group would inevitably have to go into greaterdepth on legal and policy issues.
1. DEFINITION OF TERMS
"Re-introduction": an attempt to establish a species(2) in an area which was once part of itshistorical range, but from which it has been extirpated or become extinct (3) ("Re-establishment" is asynonym, but implies that the re-introduction has been successful).
"Translocation": deliberate and mediated movement of wild individuals or populations from one partof their range to another.
"Re-inforcement/Supplementation": addition of individuals to an existing population ofconspecifics.
"Conservation/Benign Introductions": an attempt to establish a species, for the purpose ofconservation, outside its recorded distribution but within an appropriate habitat and eco-geographicalarea. This is a feasible conservation tool only when there is no remaining area left within a species'historic range.
2. AIMS AND OBJECTIVES OF RE-INTRODUCTION
a. Aims:
The principle aim of any re-introduction should be to establish a viable, free-ranging population in thewild, of a species, subspecies or race, which has become globally or locally extinct, or extirpated, inthe wild. It should be re-introduced within the species' former natural habitat and range and shouldrequire minimal long-term management.
b. Objectives:
The objectives of a re-introduction may include: to enhance the long-term survival of a species; tore-establish a keystone species (in the ecological or cultural sense) in an ecosystem; to maintainand/or restore natural biodiversity; to provide long-term economic benefits to the local and/or nataionaleconomy; to promote conservation awareness; or a combination of these.
3. MULTIDISCIPLINARY APPROACH
A re-introduction requires a multidisciplinary approach involving a team of persons drawn from avariety of backgrounds. As well as government personnel, they may include persons fromgovernmental natural resource management agencies; non-governmental organisations; fundingbodies; universities; veterinary institutions; zoos (and private animal breeders) and/or botanicgardens, with a full range of suitable expertise. Team leaders should be responsible for coordinationbetween the various bodies and provision should be made for publicity and public education about theproject.
4. PRE-PROJECT ACTIVITIES
4a. BIOLOGICAL
(i) Feasibility study and background research
An assessment should be made of the taxonomic status of individuals to be re-introduced. They should preferably be of the same subspecies or race as those which were extirpated, unless adequate numbers are not available. An investigation of historical information about the loss and fate of individuals from the re-introduction area, as well as molecular genetic studies, should be undertaken in case of doubt as to individuals' taxonomic status. A study of genetic variation within and between populations of this and related taxa can also be helpful. Special care is needed when the population has long been extinct. Detailed studies should be made of the status and biology of wild populations(if they exist) to determine the species' critical needs. For animals, this would include descriptions of habitat preferences, intraspecific variation and adaptations to local ecological conditions, social behaviour, group composition, home range size, shelter and food requirements, foraging and feeding behaviour, predators and diseases. For migratory species, studies should include the potential migratory areas. For plants, it would include biotic and abiotic habitat requirements, dispersal mechanisms, reproductive biology, symbiotic relationships (e.g. with mycorrhizae, pollinators), insect pests and diseases. Overall, a firm knowledge of the natural history of the species in question is crucial to the entire re-introduction scheme. The species, if any, that has filled the void created by the loss of the species concerned, should be determined; an understanding of the effect the re-introduced species will have on the ecosystem is important for ascertaining the success of the re-introduced population. The build-up of the released population should be modelled under various sets of conditions, in order to specify the optimal number and composition of individuals to be released per year and the numbers of years necessary to promote establishment of a viable population. A Population and Habitat Viability Analysis will aid in identifying significant environmental and population variables and assessing their potential interactions, which would guide long-term population management.
(ii) Previous Re-introductions
Thorough research into previous re-introductions of the same or similar species and wide-ranging contacts with persons having relevant expertise should be conducted prior to and while developing re-introduction protocol.
(iii) Choice of release site and type
Site should be within the historic range of the species. For an initial re-inforcement there should be few remnant wild individuals. For a re-introduction, there should be no remnant population to prevent disease spread, social disruption and introduction of alien genes. In some circumstances, a re-introduction or re-inforcement may have to be made into an area which is fenced or otherwise delimited, but it should be within the species' former natural habitat and range. A conservation/ benign introduction should be undertaken only as a last resort when no opportunities for re-introduction into the original site or range exist and only when a significant contribution to the conservation of the species will result. The re-introduction area should have assured, long-term protection (whether formal or otherwise).
(iv) Evaluation of re-introduction site
Availability of suitable habitat: re-introductions should only take place where the habitat and landscape requirements of the species are satisfied, and likely to be sustained for the for-seeable future. The possibility of natural habitat change since extirpation must be considered. Likewise, a change in the legal/ political or cultural environment since species extirpation needs to be ascertained and evaluated as a possible constraint. The area should have sufficient carrying capacity to sustain growth of the re-introduced population and support a viable (self-sustaining) population in the long run. Identification and elimination, or reduction to a sufficient level, of previous causes of decline: could include disease; over-hunting; over-collection; pollution; poisoning; competition with or predation by introduced species; habitat loss; adverse effects of earlier research or management programmes; competition with domestic livestock, which may be seasonal. Where the release site has undergone substantial degradation caused by human activity, a habitat restoration programme should be initiated before the re-introduction is carried out.
(v) Availability of suitable release stock
It is desirable that source animals come from wild populations. If there is a choice of wild populations to supply founder stock for translocation, the source population should ideally be closely related genetically to the original native stock and show similar ecological characteristics (morphology, physiology, behaviour, habitat preference) to the original sub-population. Removal of individuals for re-introduction must not endanger the captive stock population or the wild source population. Stock must be guaranteed available on a regular and predictable basis, meeting specifications of the project protocol. Individuals should only be removed from a wild population after the effects of translocation on the donor population have been assessed, and after it is guaranteed that these effects will not be negative. If captive or artificially propagated stock is to be used, it must be from a population which has been soundly managed both demographically and genetically, according to the principles of contemporary conservation biology. Re-introductions should not be carried out merely because captive stocks exist, nor solely as a means of disposing of surplus stock. Prospective release stock, including stock that is a gift between governments, must be subjected to a thorough veterinary screening process before shipment from original source. Any animals found to be infected or which test positive for non-endemic or contagious pathogens with a potential impact on population levels, must be removed from the consignment, and the uninfected, negative remainder must be placed in strict quarantine for a suitable period before retest. If clear after retesting, the animals may be placed for shipment. Since infection with serious disease can be acquired during shipment, especially if this is intercontinental, great care must be taken to minimize this risk. Stock must meet all health regulations prescribed by the veterinary authorities of the recipient country and adequate provisions must be made for quarantine if necessary.
(vi) Release of captive stock
Most species of mammal and birds rely heavily on individual experience and learning as juveniles for their survival; they should be given the opportunity to acquire the necessary information to enable survival in the wild, through training in their captive environment; a captive bred individual's probability of survival should approximate that of a wild counterpart. Care should be taken to ensure that potentially dangerous captive bred animals (such as large carnivores or primates) are not so confident in the presence of humans that they might be a danger to local inhabitants and/or their livestock.
4b. SOCIO-ECONOMIC AND LEGAL REQUIREMENTS
Re-introductions are generally long-term projects that require the commitment of long-term financial and political support. Socio-economic studies should be made to assess impacts, costs and benefits of the re-introduction programme to local human populations. A thorough assessment of attitudes of local people to the proposed project is necessary to ensure long term protection of the re-introduced population, especially if the cause of species' decline was due to human factors (e.g. over-hunting, over-collection, loss or alteration of habitat). The programme should be fully understood, accepted and supported by local communities. Where the security of the re-introduced population is at risk from human activities, measures should be taken to minimise these in the re-introduction area. If these measures are inadequate, the re-introduction should be abandoned or alternative release areas sought. The policy of the country to re-introductions and to the species concerned should be assessed. This might include checking existing provincial, national and international legislation and regulations, and provision of new measures and required permits as necessary. Re-introduction must take place with the full permission and involvement of all relevant government agencies of the recipient or host country. This is particularly important in re-introductions in border areas, or involving more than one state or when a re-introduced population can expand into other states, provinces or territories. If the species poses potential risk to life or property, these risks should be minimised and adequate provision made for compensation where necessary; where all other solutions fail, removal or destruction of the released individual should be considered. In the case of migratory/mobile species, provisions should be made for crossing of international/state boundaries.
5. PLANNING, PREPARATION AND RELEASE STAGES
Approval of relevant government agencies and land owners, and coordination with national and international conservation organizations. Construction of a multidisciplinary team with access to expert technical advice for all phases of the programme. Identification of short- and long-term success indicators and prediction of programme duration, in context of agreed aims and objectives. Securing adequate funding for all programme phases. Design of pre- and post- release monitoring programme so that each re-introduction is a carefully designed experiment, with the capability to test methodology with scientifically collected data. Monitoring the health of individuals, as well as the survival, is important; intervention may be necessary if the situation proves unforseeably favourable. Appropriate health and genetic screening of release stock, including stock that is a gift between governments. Health screening of closely related species in the re-introduction area. If release stock is wild-caught, care must be taken to ensure that: a) the stock is free from infectious or contagious pathogens and parasites before shipment and b) the stock will not be exposed to vectors of disease agents which may be present at the release site (and absent at the source site) and to which it may have no acquired immunity. If vaccination prior to release, against local endemic or epidemic diseases of wild stock or domestic livestock at the release site, is deemed appropriate, this must be carried out during the "Preparation Stage" so as to allow sufficient time for the development of the required immunity. Appropriate veterinary or horticultural measures as required to ensure health of released stock throughout the programme. This is to include adequate quarantine arrangements, especially where founder stock travels far or crosses international boundaries to the release site. Development of transport plans for delivery of stock to the country and site of re-introduction, with special emphasis on ways to minimize stress on the individuals during transport. Determination of release strategy (acclimatization of release stock to release area; behavioural training - including hunting and feeding; group composition, number, release patterns and techniques; timing). Establishment of policies on interventions (see below). Development of conservation education for long-term support; professional training of individuals involved in the long-term programme; public relations through the mass media and in local community; involvement where possible of local people in the programme. The welfare of animals for release is of paramount concern through all these stages.
6. POST-RELEASE ACTIVITIES
Post release monitoring is required of all (or sample of) individuals. This most vital aspect may be by direct (e.g. tagging, telemetry) or indirect (e.g. spoor, informants) methods as suitable. Demographic, ecological and behavioural studies of released stock must be undertaken. Study of processes of long-term adaptation by individuals and the population. Collection and investigation of mortalities. Interventions (e.g. supplemental feeding; veterinary aid; horticultural aid) when necessary. Decisions for revision, rescheduling, or discontinuation of programme where necessary. Habitat protection or restoration to continue where necessary. Continuing public relations activities, including education and mass media coverage. Evaluation of cost-effectiveness and success of re- introduction techniques. Regular publications in scientific and popular literature.
Footnotes:
IUCN/ SSC CONTACT INFORMATION:
The IUCN/SSC Re-introduction Specialist Group (RSG) is a disciplinary group (as opposed to mostSSC Specialist Groups which deal with single taxonomic groups), covering a wide range of plant andanimal species. The RSG has an extensive international network, a re-introduction projects databaseand re-introduction library. The RSG publishes a bi-annual newsletter RE-INTRODUCTION NEWS.
If you are a re-introduction practitioner or interested in re-introductions please contact:Mr. Pritpal S.SooraeSenior Conservation Officer IUCN/SSC Re-introduction Specialist Group (RSG) Environmental Research & Wildlife Development Agency (ERWDA) P.O. Box 45553 Abu Dhabi United Arab Emirates (UAE)
Tel: (D/L) 971-2-693-4650 or general line: 693-4628 Fax: 971-2-681-7361 E-mail: PSoorae@erwda.gov.ae
< kahanpaa@math.jyu.fi> Apr.20.2004