For pdf versions or paper copies, take contact to the Society
Lauri Kahanpää
Dear Friend.
Three Finnish issues of the Bulletin appeared in 2011, none in 2012. This issue - number 2/2013 - contains English translations of most of the material in the Finnish Bulletin number 1/2013. The material in vol 2011 will be summarized in another English issue to appear later

Lauri Kahanpää
In 2012 all our efforts were concentrated on winning a suite of macabre legal processes, which are seriously threatening the future of credible LWfG conservation efforts in Finland. (See 4/2011). So in 2012 no issue of the Bulletin was published, not even the long promised English summary of the three 2011 Finnish issues containing a description of the current legal situation. WeÕll do our best to have this corrected soon.
In spite of the silence, the Friends have been active on the practical level all the time taking care of the only Lesser White-fronted Geese breeding in Finland Ð our own. To mark the real aims of our activities, we decided to skip all the Franz Kafka-style troubles in this issue and concentrate on birds. The unpleasant court material will come later.
It might be worthwhile mentioning from where we gather most of our data. Of course, our members are doing observations of their own, mostly abroad or observing captive birds. A wealth of information is available on the net. In Finland, BirdLife collects observations of all bird species in their database ÓTiiraÓ. Fresh observations on migration generally quickly appear on the discussion forum ÓLintuverkko. Foreign observations of LWfG are collected and published at www.piskulka.net, which should collect all observations but in practise concentrates on the birds still breeding in Norway. In particular, it presents no news concerning Swedish LWfG. To find out about the complete status of LWfG in Scandinavia one has to visit the Swedish page http://www.artportalen.se and ask oneÕs friends and neighbours. A general picture of Geese in Europe is reflected in the scientific journals, in particular in Wetlands InternationalÕs Goose Bulletin http://www.geese.org/gsg/goose_bulletin.html . To learn more about the LWfG in general one has to learn some basic Russian and read the annual journal Cazarca, distributed by us in Finland and published in Russia by the RGG also known as the GSDSG, the Goose, Swan, and Duck Study Group of Northern Eurasia. Of course correspondence with goose experts in various countries as well as our regular participation in conferences also bring in a lot of information and understanding.
Lauri Kahanpää.
The lemming years 2010 and 2011 supplied plenty of easy food for small predators in northern Norway. The Lesser White-fronted Geese could breed in peace. As a result a long time high of 13 broods with an average of 3.4 goslings each was counted in August 2010 in the Valdak marshes, the traditional the autumn migration gathering area. One year later, the return to reality was abrupt; only 3 broods with a total of 12 young appeared. Also most of the previous yearsÕ young birds were absent.
ÒOnce in four years, the lemmings living on the highest mountain tops in northern Norway begin their enormous expansion over all of LaplandÓ, That is the common belief. Siberian relatives of our lemmings may still show a regular pattern like that but in Europe predicting the lemming years is more difficult, if not impossible. According to friendly personal information from Prof. Heikki Henttonen (of the Finnish Forest Research Institute Metla), the following were lemming years 1937-1938, 1942 (local), 1946 (local), 1959 (local), 1960 (small), 1969Ð1970 (great), 1974, 1978 (intermediate), 1982 (weak), 1997-1998 (ended prematurely), 2001 (weak), 2007 (focus in Norway), and 2010-2011(strong). The distances between the maximum years are 4, 4, 23-24, 4, 4, 4, 15, 4, 6, and 3-4 years. If there is double periodicity, we should now expect another long gap. Since the time line above only contains two of the long gaps, I would not yet bet on anything. Instead, I have for much more than a decade been engaged in attempts to forecast the numbers of Lesser White-fronted Geese. At first sight the recent sudden temporary increase and crash of the Norwegian geese seem to be in conflict with my earlier expectations. But do they? And what is the connection to the lemmings? Let us see.
Here are the observations:
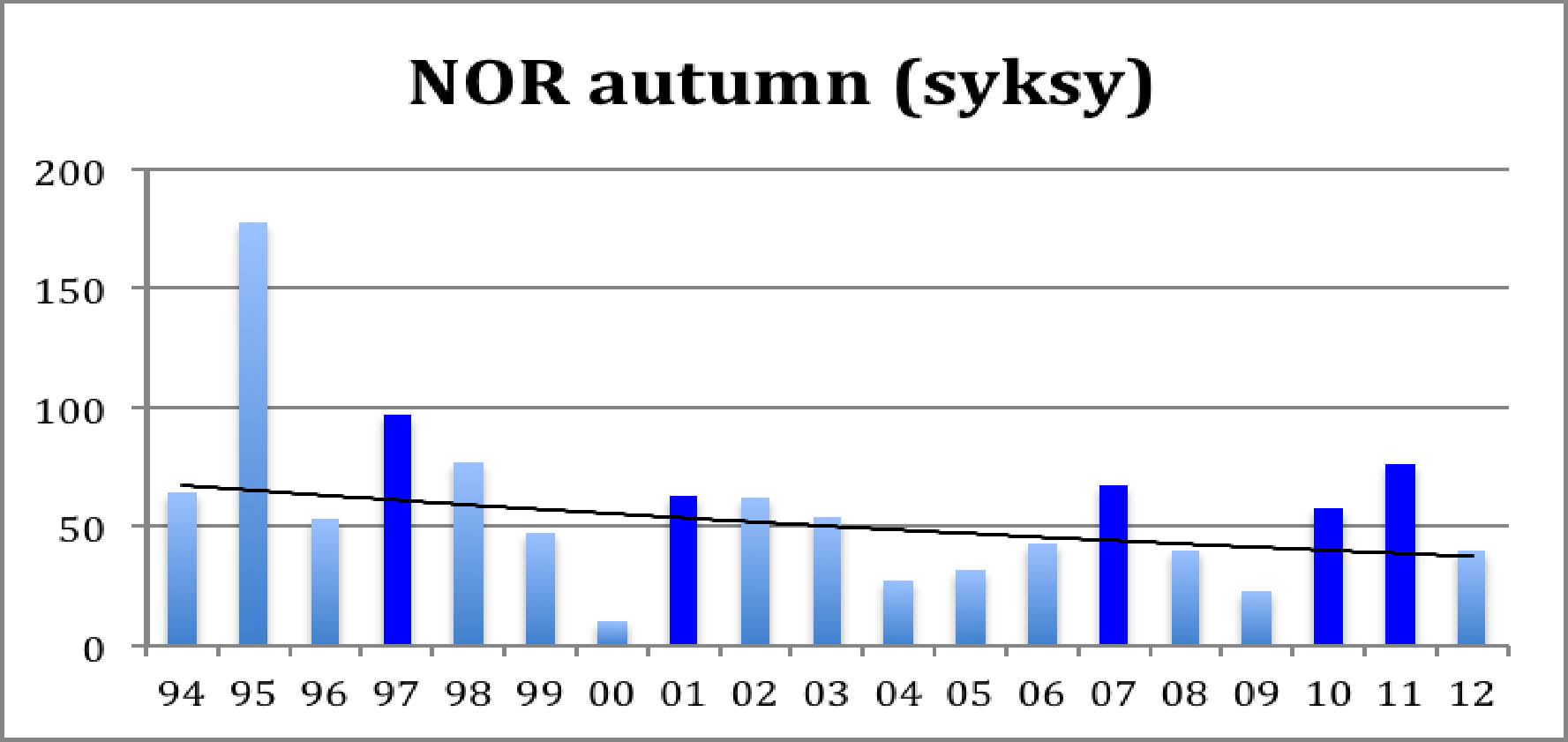
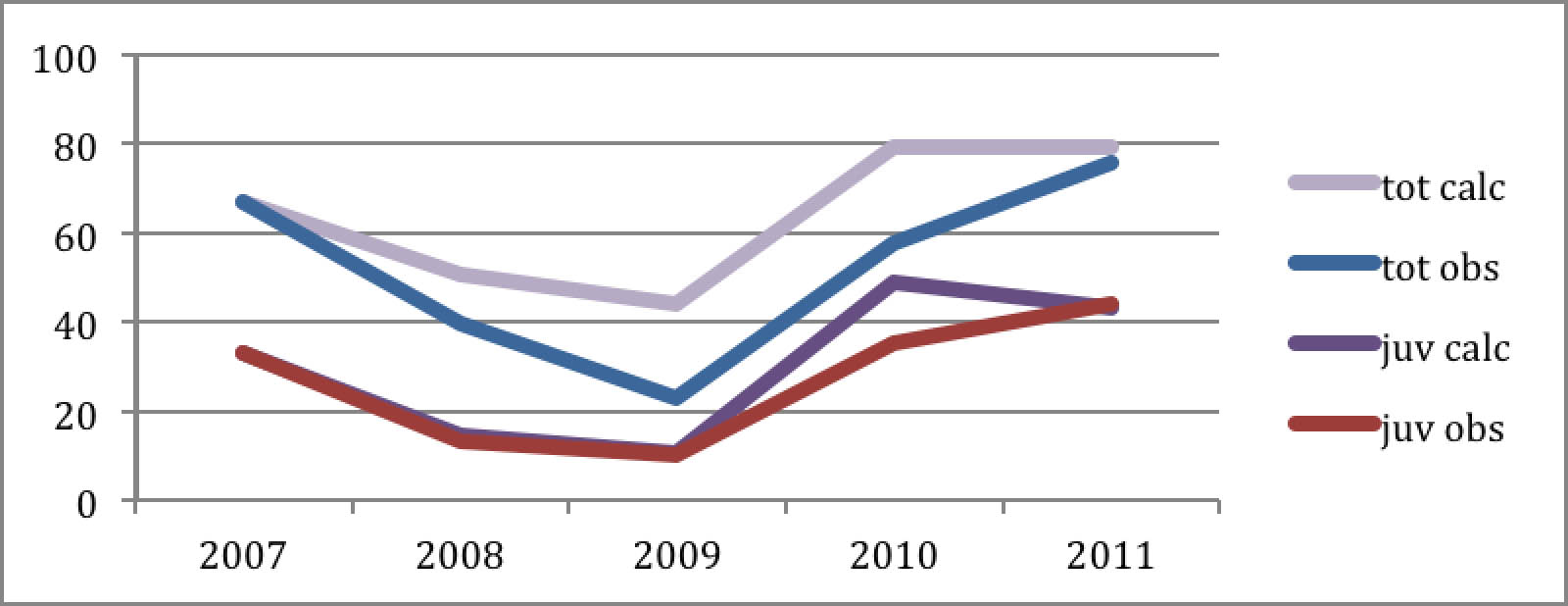
Diagram 1: Autumn LWfG in Norway (ad + juv)
Diagram 1 represents the "Piskulka page" data of August concentrations of Lesser White-fronted Geese in the Valdak marshes at Porsanger Fiord Norway. Lemming years are marked dark. The down sloping curve is the exponential trendline.
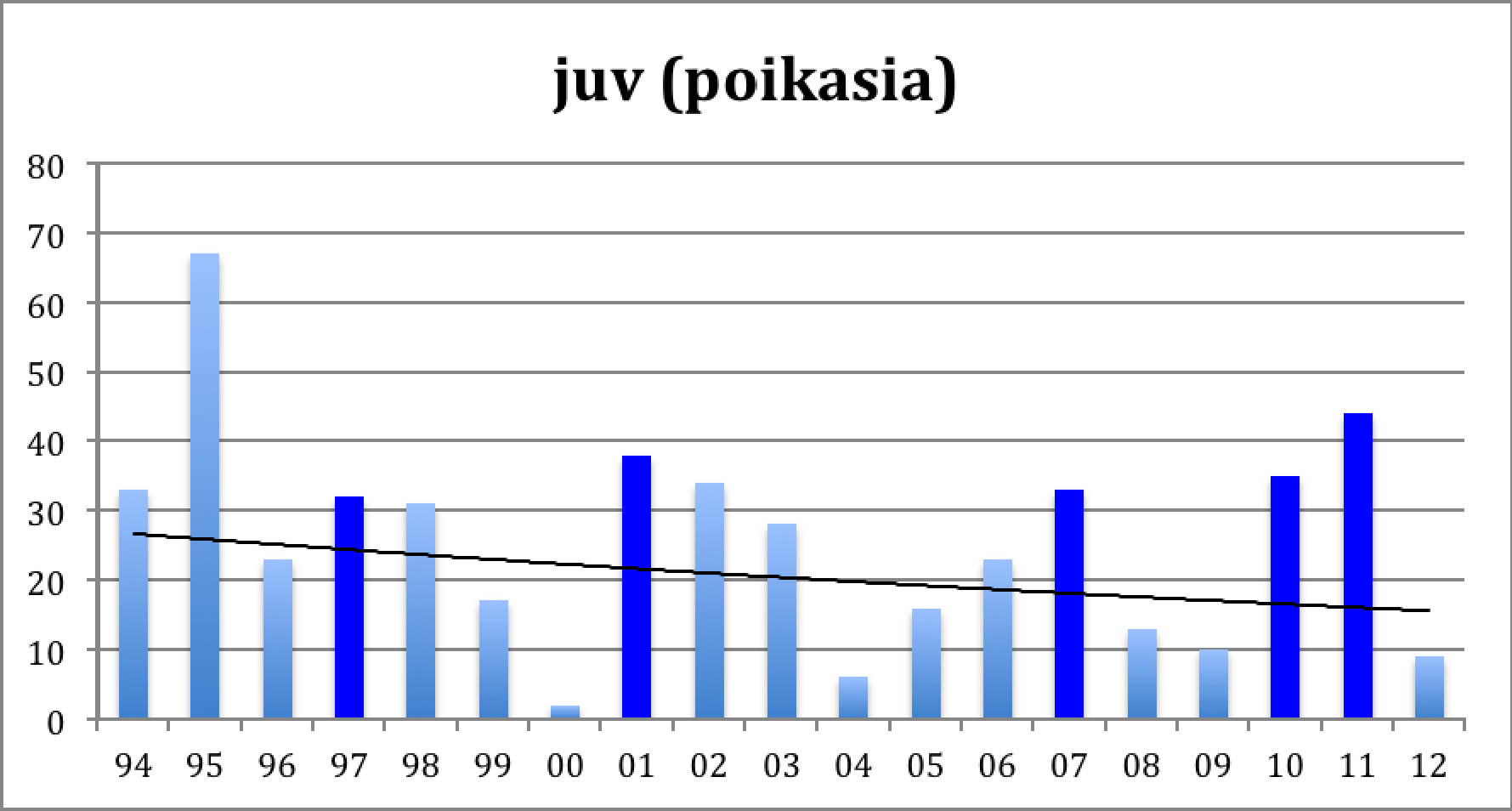
Diagram 2: Goslins
Diagram 2 shows the "Piskulka page" numbers of goslings in August at Valdak. Lemming years are again marked dark, the curve represents the trend as an exponential decrease. .
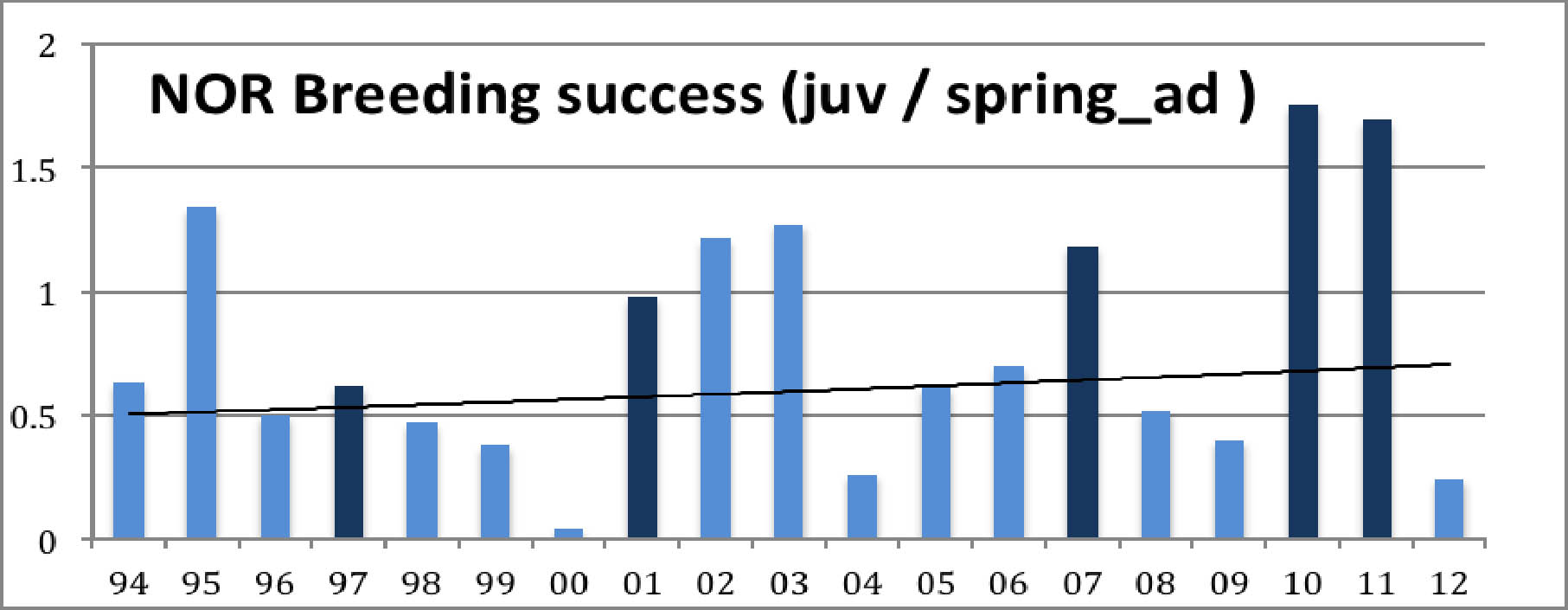
The year 2000 had unusually adverse weather conditions. Apart from that, both diagrams show a decrease of the population with peaks at the lemming years. Diagram 1 shows an average decrease of the total autumn population by 3.0 % / year which corresponds to about halving the total population in 20 years. Diagram 2 shows an average decrease of the autumn juveniles population by 2.7 % / year. The fact that the juveniles have a slightly smaller decrease rate than the overall population reflects a slight increase in the average breeding result. This is confirmed by calculating the breeding result (autumn juv / spring adult). And yes, there is a slight increase in the average result: from 0.68 in the first decade to 0.77 in the second, 0.71 over the whole period. For more detail, let us map the results year by year:
Diagram 3: Breeding result
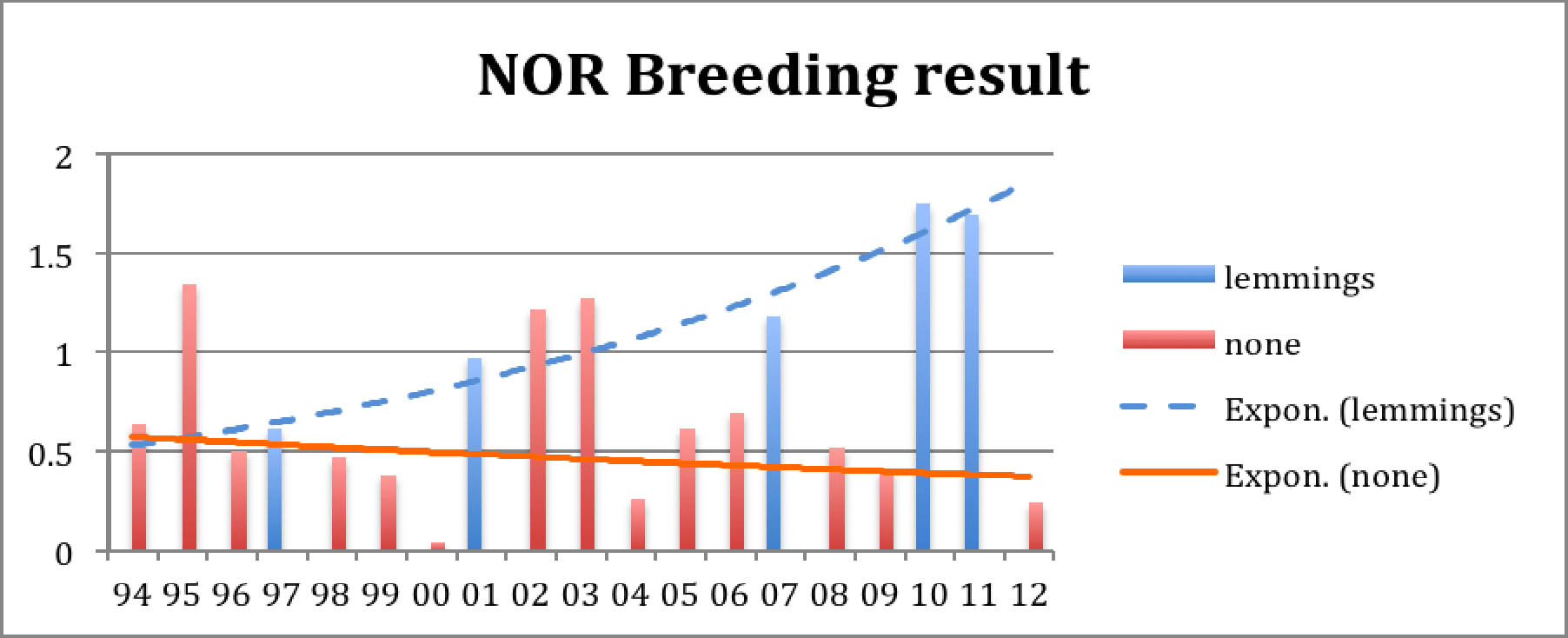
What strikes the eye is the extreme year 2000 and Ð more interesting - the great difference between lemming years and other years. Also, their difference seems to be increasing, and the drop after a lemming year seems to become more dramatic. Also the breeding success in normal years seems to decrease while the breeding result in lemming years becomes better. Let us illustrate this by adding trend lines for both::
Diagram 4: Breeding result in lemming and non-lemming years
The interaction
In lemming years predators find so much easy prey that they loose interest in hunting the geese. This wellknown fact explains the good breeding result. Like the geese, also the predators multiply in the lemming year, so after the crash of the rodent population there are many predators around, and they are hungry. This explains the sudden drop in the goose breeding result in the next year. A gradual recovery follows when the predators normalize. This cycle is clearly visible in both diagrams, Diagram 1 shows that after the crash it takes about three or four years for the goose population to shrink to about half of the previous peak size. Diagrams 2 and 3 give the explanation: very few juvenils are added. Adult and subadult mortality are probably slightly increased in the post-lemming summer and migration/winter mortality goes on as usual.
The apparent intensification of the breeding result cycle might have to do with an overall increase trend in small predators, in particular the red fox, in the area. Also, changes in observation intensity/skill and reporting are possible over the time span of two decades.
Interpretation
We should keep in mind how the Lesser White-fronted Geese's mortality depends on their migration and wintering pattern. The basic parameters are the age of the bird and the migration route chosen. Generally young birds have a much higher mortality than adults, and birds taking the long eastern route have a higher mortality than birds migrating more directly to Greece.
Let us make this explicit and simple: Let us adopt the WWF mortality parameters from the late 1990:s and beginning 2000:s: The mortality during the first wintering is 78 % for goslings and 16% for adult birds. Let us apply this to understanding a lemming four year cycle. We take as initial data the result of the lemming year 2007: 29 adults, 5 subadults and 33 goslings in autumn. After one migration round, in spring 2008 we would have remaining 24 adults, 4 subadults and 7 juvenils. In the post lemming year they will have a bad breeding result. To be exact, the 2008 result was 0.52 young/(sub)adult. For simplicity, summer mortality is included in the mortality numbers already, so we can say all spring birds survive in autumn, and we arrive at an autumn 2008 flock of 51 birds, namely 29 adults, 7 subadults and 15 juvenils. We can continue the same way, and compare with observations. In the post-lemming years 2008-2009 the breeding results were 0.52 and 0.36, in the two following lemming years they were 1.75 and 1.69. Adopting these numbers and the old mortality parameters from ten years ago we calculate the following results for the autumn flock, total and juveniles:
Diagram 5: Observation vs. calculation of over the lemming cycle 2007-2011
The calculated curves are similar to the observed both in their general shape reflecting the dynamics of the lemming years and in the overall result over this period from one lemming year to the next. As the reproduction parameters were taken directly from observations, this indicates that the mortality parameters were correct: They have not changed since ten years ago.
The need for more details
In the data, subadults are not always counted separately from adults. Therefore, in the above calculations subadult birds are mostly treated as adults, in particular the breeding index takes this correctly into account. But subadult birds not only are not breeding. They also prefer the eastern much more dangerous migration route. Similarly, non-breeding adults prefer the eastern migration route. The differences in mortalities cannot be calculated from the current data. The mortality 16% for all ÒadultsÓ actually is an average for all adults and subadults. It is optimistic in the post-lemming years when there are many subadults and non-breeding or failed adults around. The opposite takes place in lemming years. This phenomenon magnifies the lemming year fluctuations in goose numbers but has only a small effect on their overall trend.
Today
Today is May 10, 2013 and the Lesser White-fronted Geese are just returning to Norway. On their spring migration they have been counted in Greece and Hungary. Just like expected, mortality was larger than average. In Greece the largest flocks contained 75 geese in the previous winter, now 22 birds (29%) less. In Hungary, the drop is even bigger (40-48 %).
Predictions and observations
I must admit that I did not think of the lemmings at the Goose 2001 in Roosta when I presented the first predictions for the future of the Norwegian Lesser White-fronted Geese. The prediction built solely on the average breeding results and adult / juv mortalities which were estimated carefully. The main result was an explanation of the observed average annual decrease of about 5%. Three years later, at the a meeting an improved model took into account random changes in the parameters. Mathematically that was interesting but the only substantial change in the predictions was quantitative version of the fact that large oscillations in annual mortality increase the risk for final extinction of a small population. The update at the last public scientific Lesser White-fronted Goose meeting Xanten in 2007 brought no changes but an extension of the simple prediction scheme to a large model encompassing not only the Norwegian geese but also the Swedish, Russian, captive and possible future Finnish populations and their natural and artificial interactions like transporting geese from one location to another. For future revisions of the parameters, they were treated as inputs, not parts of the model structure. For details, see the conference proceedings (Vogelwelt---) The full model is available at www.piskulkaconf.tk by clicking Ó Calculate easily the effects of protection measures on LWfG populations! Ó and Background document Ó . Using the model it is easy to check that the recent peak in Norwegian Lesser White-fronted Goose numbers is entirely due to the lemming years.
A final remark
A quick check in data on the Swedish reintroduced population revealed a similar effect. The connection between rodents, predators and geese is well known but I was truly surprised by its strength. The effect of climate change on lemming years may prove very important for the future of the geese, but for the time being a careful look at the observations has confirmed the importance of captive breeding and quick and strong reintroduction programs not only in Sweden.
Thomas Heinicke and Lauri Kahanp (ed.)

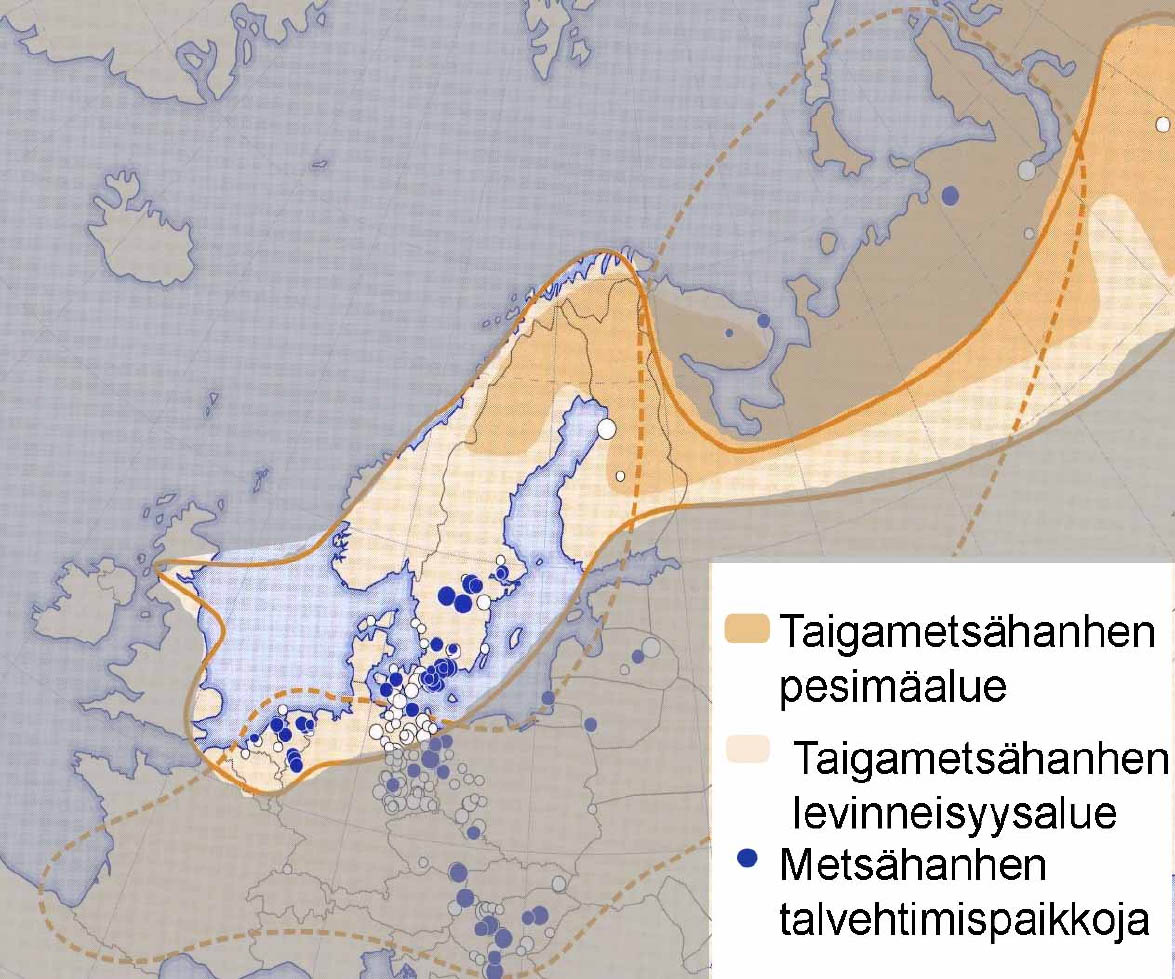
In contrast to the Lesser White-fronted Goose, the Bean Goose has several subspecies. The ones breeding in Finland are Taiga Bean Geese (Anser f. fabalis), but also large numbers of Tundra Bean Geese (Anser f. rossicus) migrate through SW Finland. These subspecies are sufficiently independent to be considered separate conservation units. Recent observations indicate that Finland is the key country in protecting the Taiga Bean Goose (Anser f. fabalis) Ð now globally threatened.
He scientific content and illustrations of this paper is completely based on a talk given by Thomas Heinicke at the GOOSE 2011-conference, and is publiched on his request to draw attention to the special status of the Taiga bean Goose in Finland
The Taiga Bean Goose (Anser f. fabalis) is almost the size of the Greylag Goose. Its beak is long and slim, usually largely orange in colour, but sometimes dark more than halfway to the tip much like the beak of the Tundra Bean Goose (Anser f. rossicus), whose beak is always dark and relatively thick at the base. The Tundra Bean Goose is smaller, has a darker head and shorter neck. For exact identification of the subspecies, see http://www.birdlife.fi/suojelu/lajit/tundrametsahanhi.pdf .
The Bean Goose is a common bird in Europe but the subspecies (Anser f. fabalis) only breeds on a strip extending from Sweden over Finland and Russia to the Urals. Their numbers, in particular the proportions of the subspecies are counted in the wintering areas in western Europe. Some Bean Goose migrate to Asia, but not the ones discussed here.
During the last decade the Bean goose subspecies were counted separately. The alarming observation is a clear decline in the numbers of the Taiga subspecies, ie. the ones breeding in Finland. The census results are listed below:
Tammikuu
S
DK
D
PL
UK
Yht.hav
Arvio
2004
9.326
10.683
35.000
3.800
375
69.200
70.000-90.000
2005
34.560
8.728
42.000
490
418
86.200
70.000-90.000
2006
19.289
16.279
52.000
1.500
469
89.500
70.000-90.000
2009
32.500
13.836
22.500
1.500
471
70.800
60.000-65.000
2011
8.201
20.000
12.100
1.790
453
42.544
45.000
individuals in just two years (2009-2011)! What can cause this? Are there problems with the breeding or mortality? Breeding success can be monitored by counting the percentages of young birds in the autumn flocks. This was done in central Sweden in 2009 and repeated in 2010. Unfortunately, the subspecies were not counted separately. Here are the results:
Aika
nuoria
yhteensä
poikuekoko
poikueita
Syyskuu 2009
22,9%
6.710
2,56
209
Lokakuu 2009
20,90%
3.727
Syyskuu 2010
26,1%
4.202
Lokakuu 2010
17,2%
1.538
2,49
160
Marraskuu 2010
7,7%
2.521
In early autumn there were quite high juvenile percentages, even somewhat larger than in arctic geese. In late autumn there were obviously lower juvenile percentages. This can be a result of differences in migration pattern of successful and unsuccessful breeders or an involvement of different subpopulations: In Sep+Oct there could be a large proportion of local Scandinavian breeders. Later Russian breeders would dominate. These may have worse breeding results or have lost some juvenils during their migration, possibly in Finland. To find out more one can look at neck banding data from northern Sweden and Germany:
Rengastusvuosi
lkm
% elossa 2006/2007
% elossa 2007/2008
% elossa 2008/2009
% elossa 2009/2010
% elossa 2010/2011
Kevät 2007
1
100
100
100
100
100
Kevät 2008
27
100
85,2
77,8
51,9
Kevät 2009
11
100
72,7
45,5
Kuolleisuus Ruotsissa
14,3 (4/28)
14,3(5/35)
33,3(10/30)
Saksa (Oderin alajuoksu) syksy 2007
33
100
57,6
33,3
12,1
Kuolleisuus Saksassa
42,4
42,1
63,6
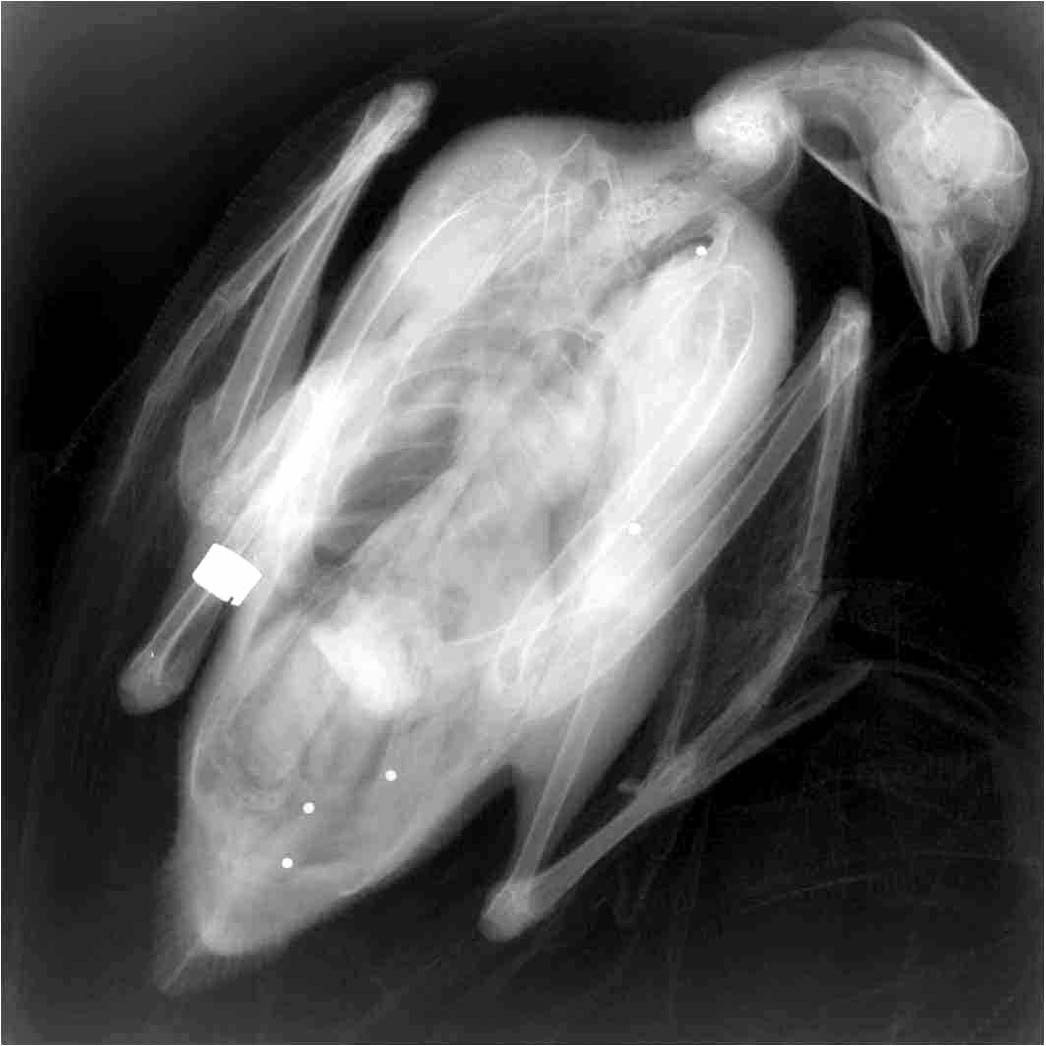
One observes the almost double mortality rate of German Bean Geese in comparison to the north Swedish data. The observed populations represent different breeding areas and different hunting pressure. The hunting pressure may be estimated by finding out how many birds carry lead pellets from shooting. They can be seen in x-rays (5 pellets in the picture below).
The following table displays the numbers and percentages of Geese caught in eastern Germany:
nuoria puhtaita
nuoria haulein
aikiuisia puhtaita
aikuisia haulein
haulein yht
merihanhi
1
0=0%
58
14=19,4%
19,2
tundrahanhi
35
0=0%
73
20=21,5%
15,6
tundra-metsähanhi
30
0=0%
44
24=35,3%
24,5
taiga-metsähanhi
4
0=0%
13
11=45,8%
39,3
21,1
Clearly, the worst stricken Geese were the Taiga Bean Geese. An important observation is that no juv birds of any species carried any lead pellets. This could be explained by a possible absence of significant hunting pressure before arriving in Germany. To check this hypotheses, one may look at the hunting bag statistics in the relevant countries (cit. Hirschbach & Heyd 2005). hunting bag year remarks
saalis
tilastovuosi
Huom
Norja
0
rauhoitettu
Suomi
7.900
2009
Pääosin taiga-
Ruotsi
3.450
2005/2006
Pääosin taiga-
Tanska
886
2005/2006
Pääosin taiga-
Saksa
4.255
2005/2006
Pääosin tundra 300-500 taiga
Puola
13.812
2005/2006
Pääosin tundra 200-400 taiga
Baltian maat
1.127
2005/2006
Pääosin tundra 100-200 taiga
V-venäjäja Ukraina
?
Pääosin tundra 200-400 taiga
Venäjä
5.000-10.000
Vähintään
Yhteensä
35.000-40.000
The subspecies separation is not precise enough for final conclusions but probably the annual bag contains some 15,000 to 20,000 Taiga Bean Geese, half of which are shot in the territory of the European Union.
So the Taiga Bean Goose is globally threatened. It clearly meets three IUCN Red List criteria (v 3.1, 2001):
- population size reduction of more than 30 % over the last 10 years or 3 generations (A2)
- population size reduction of more than 30 % projected or suspected to be met within the next 10 years or 3 generations (A3)
- population size reduction of more than 30 % over any 10 year or 3 generation period (time period include past+future), where reduction or its causes may not have ceased (A4)
To sum up, this is what we know about the Taiga Bean Goose
- Recently there has been a strong population decline in wintering Taiga Bean Geese in Europe (mostly birds of Russian breeding origin)
- A complete population crash will happen within the next 5-10 years. (Actually there has been a net loss of up to 10,000 birds per year!)
- The Taiga Bean Goose now qualifies as a globally threatened species. Urgent international and national actions are needed to protect them against extinction.
- Most relevant factors for the decline are overhunting (breeding, staging and wintering areas) and habitat loss (breeding areas, e.g. Western Siberia & Finland)
At our Goose Farm in Hmeenkoski, Finland we also have some Bean Geese. If things continue the way they are going, the Friends of the Lesser White-fronted Goose might end up re-introducing two Goose species in Finland! Please, help to protect the Taiga Bean Goose!
Extinct birds:
Lauri Kahanpää
The class of birds, Aves, exists and produces new species since about 100 million years. Most of them have gone extinct, of course, but right now we are witnessing an unusually abrupt wave of simultaneous extinctions: since the year 1500 more than 200 bird species have disappeared forever Ð on average more than one species in three years At least 91 known species have died since 1681. A list of these and the most critically endangered species is printed in grey on the outer and inner cover pages of his issue of the LWfG . (Source: Ornithology.com Home) No one of these species went extinct in Europe. We Finns share the responsibility for seeing to it that the Lesser White-fronted Goose will not be the first species to break this trend.
For a long time the famous Solnhofen Archeaopteryx was the only known fossil bird. The Age of the Dinosaurs, the Mesozoic era, consists of the Triassic, Jurassic and Cretaceous eras. The Triassic began 252 million years ago in the wake of the PermianÐTriassic extinction event, the largest well-documented mass extinction in Earth's history, and the Cretaceous ended 66 million years ago with another mass extinction which is known for having killed off the non-avian dinosaurs. The Solnhofen limestone was formed towards the end of the Jurassic, about 155 milion years ago. Recent fossil discoveries have confirmed the Jurassic origin of birds. One of these findings, the Confuciusornis, is depicted here. In contrast to the equally old Archeaopteryx, the beak of Confuciusornis has no teeth. So birds are not only descendants but were also contempo-raries of the dino- and pterosasurs. .
From the Cretaceous era already many bird orders are known. These include not only extinct orders like Enantiornithes, Hesperornithi-ormes, and Ichthyornithiformes but also some of the still existing ones. An ornithologist in the Cretaceous era could well have observed ancestors of modern birds (Neornithes), among them Shorebirds/Gulls as well as Ducks/Geese. .
After having survived the CretaceousÐTertiary extinction event, most probably caused by the the Chicxulub Yukatan asteroid impact modern birds, much likemammals, entered a period of diversification filling ecological niches left empty by extinction of other animals, in particular archaic birds and non-avian dinosaurs. In the first Cenozoic epoch, the Paleocene, large flightless birds appeared, including the Gastornis in Europe and North America, and terror birds in South America which survived until the Pleistocene, almost modern times. In the late Paleocene, early owl types appeared. By the Oligocene, 30 million years ago, most modern bird types had appeared including cranes, hawks, pelicans, herons, owls, ducks, pigeons, loons, woodpeckers and even perching birds, the most modern order, today encompassing about half of all bird species. In spite of this, the EarthÕs bird fauna still was different from what we observe today, and has gone through many changes during the remaining 30 million years. Their evolution has been directed by natural factors such as climate changes and competition by other species. An example of the latter seems to be an extinction wave of aquatic birds before the Ice Age by competition of sea mammals. Also natural catastrophes like volcanic eruptions and comet and asteroid impacts have caused high extinction rates, at least locally. Of course, this still goes on today, in particular by the climate changes associated to the various phases of the Ice Ages. Fossils of more than 300 extinct bird species are known and named from the last one million years and there must be much more since bird bones are fragile and fossilize rarely Ð in particular complete skeletons are extremely difficult to find.
All the time, diversification has gone on producing new species. An easily recognisable phenomenon is the formation of species pairs like the Greater and Lesser White-fronted Goose during the last icing period. Since then, also the impact of humans on birds becomes important. Well known is the extinction of the New Zealand giant Moas already by the Maoris, but more generally the impact of early man on the bird fauna is largely unknown. In contrast to that there exists a rather complete scientific record of the last 500 years. The record is embarrassing. About 200 bird species have gone extinct. What is going on is an extremely rapid wave of mass extinctions doubtlessly caused by humans in many ways. Remembering the excessive hunting pressure on the Lesser White-fronted Goose we first think of direct killing. And yes, both the Dodo and the Great Auk were hunted to extinction. More subtly, also the Passenger Pigeon was hunted to death Ð it was specialized to living in huge swarms and could not recover after moderately looking hunting. Today the absence of sufficiently large swarms may affect the Lesser White-fronted Goose as well. Besides direct hunting, many human activities affect the bird fauna. Agriculture, forestry, drying of swamps, building water reservoirs and other land use has greatly changed the environment. The effects of anthropogenic climate change can also already be seen. In fact birds, being easy to observe and able to change locality very quickly, are some of the most sensitive indicators of climate change. This also applies to the Lesser White-fronted Goose, via the Lemmings!
There is one more way in which humans have caused extensive damage to the avian fauna. In fact most extinctions in historical time have been caused by the introduction of invasive foreign species. The classical example is the cat on an isolated lighthouse island that both found and killed a new flightless bird species. I have not verified this story but regardless of its truth it illustrates the mechanism behind most bird species extinctions. Most of them did happen in isolated areas: 30 % of them actually lived on the Hawaii islands and 10 of the 13 endemic bird species on the Guam island were killed in 30 years after the introduction of the Brown Tree Snake. By definition, a foreign species is a species occurring, as a result of human activities, beyond its normal distribution. A foreign species is called invasive if it causes damage threatening environmental, agricultural or other social resources. Usually, foreign species are not introduced on purpose - the opening of the Suez Canal connected two water bodies leading to extensive exchange of species between the Mediterranean and Red Seas. On the other hand species like the Barnacle Geese in Finland cannot be called a foreign or invasive species since they have always migrated over the country and had the opportunity to stay and breed here.
No European species appears on the list of recently extinct bird species printed as background on the cover pages of the Bulletin. It is easy to guess why: already before the year 1500, the effect of human activities on our densely inhabited continent was so strong that most damage was done before being registered. This does not mean that new extinctions are impossible. On the contrary, 68 of the 524 European bird species are classified as endangered. This is 13 %, slightly more than the global 12 %. Most threatened is the Slender-billed Curlew (Numenius tenuirostris). Of course, also the Lesser White-fronted Goose is on the list. It is classified as vulnerable (VU) on the scale:
- Not Evaluated (NE)
- Data Deficient (DD)
- Extinct (RE)
- Extinct in the Wild (EW)
- Critically endangered(CR)
- Endangered (EN)
- Vulnerable (VU)
- Near threatened (NT)
- Least Concern (LC)
If the current trend continues, it will not take long before the Lesser White-fronted Goose must be classified as (globally) endangered and RE (Regionally extinct). If breeding in captivity continues to be only a hobby for private persons, it is possible that the Lesser White-fronted Goose will become the first European bird species to become extinct in historical time. To prevent that from happening is the main and only objective of us, the Friends of the Lesser White-fronted Goose.
ISSN-L 1798-0437
ISSN 1798-0437
< lauri.v.kahanpaa@jyu.fi> Technical update done Dec 18 2014