Lauri Kahanpää
Vuonna 2012 ei ilmestynyt yhtään Kiljuhanhen Ystävä -lehteä, Viime vuoden kaikki voimavarat keskitettiin nimittäin oikeustaisteluun Suomen kiljuhanhien puolesta. Vuoden 2011 lehdistä voi lukea, mistä on kysymys.
Kiljuhanhen Ystävät ry on kuitenkin toiminut ja toimii edelleen tehokkaasti myös käytännössä lajin pelastamiseksi. Siksi tämä vuoden 2013 ensimmäinen lehti sisältääkin pelkästään lintujuttuja. Samoin varmaan seuraava numero. Pykälien vuoro tulee taas aikanaan.
Kiljuhanhitietoa kertyy oman toimintamme ja jäsentemme havaintoretkien lisäksi muutamasta muusta lähteestä, joista osaa lukija voi käyttää itsekin. Suomessa BirdLifen "Tiira"-havaintopankki ja "Lintuverkko" -keskustelupostilista raportoivat joistakin lähinnä muuttoon liittyvistä havainnoista. Tietoja muista maista julkaisee ennen kaikkea norjalais-suomalainen kiljuhanhisivu www.piskulka.net, joka tosin keskittyy ensisijaisesti Norjassa pesiviin kiljuhanhiin ja sensuroi Ruotsin kiljuhanhikantaa ja tarhakantoja koskevat tiedot. Niitä tietoja joudumme hakemaan Artportalenista http://www.artportalen.se ja kysymään tutuilta Ruotsista ja Saksasta. Perusteellisempia tieteellisiä kirjoituksia on luonnollisesti alan lehdissä, etenkin Wetlands Internationalin hanhitutkijoiden internetissä julkaistamassa Goose Bulletinissa http://www.geese.org/gsg/goose_bulletin.html ja venäjällä vuosittain ilmestyvässä Cazarca-lehdessä, jonka jakelua Suomessa olemme hoidelleet monta vuotta. Kumpikin julkaisusarja on esitelty Kiljuhanhen Ystävässä. Tärkeää on tietenkin myös kirjeenvaihto ulkomaisten hanhiasiantuntijoiden kanssa sekä osallistuminen alan konferensseihin, joista olemme yleensä raportoineet tässä lehdessä.
Lauri Kahanpää.
Sopulivuosina 2010 ja 2011 pienpedoilla riitti syötävää Ruijassa. Kiljuhanhet saivat pesiä rauhassa. Niinpä vuonna 2011 laskettiin 13 poikuetta, joissa keskimäärin 3,4 poikasta. Paluu arkeen oli karu: seuraavana vuonna tuli syysmuutto-kerääntymälle vain 3 poikuetta, joissa yhteensä 12 poikasta. Edellisen vuoden nuoret linnutkin olivat hävinneet jonnekin. Tämä ilmiö antoi aiheen tarkastella sopulivuosien ja kiljuhanhien välistä yhteyttä laajemminkin. Tulos on yllättävä: Muuttoreittien suojelu ei ole vaikuttanut mitään Norjan kiljuhahien määrään: niiden tilapäiset runsastumiset &endash; etenkin viimeisin - ovat aiheutuneet pelkästään sopulivuosista.
Sopulivuosien ennustaminen on tunnetusti vaikeaa, jopa mahdotonta. Niinpä en aio sitä tässä yrittääkään, mutta olen pitempään kuin vuosikymmenen ajan yrittänyt ennustaa kiljuhanhien lukumäärän kehitystä. Onko tapahtunut kehitys ja Norjan kiljuhanhien lukumääräpiikki sopusoinnussa ennusteen kanssa? Katsotaanpa.
Aluksi havainnot:
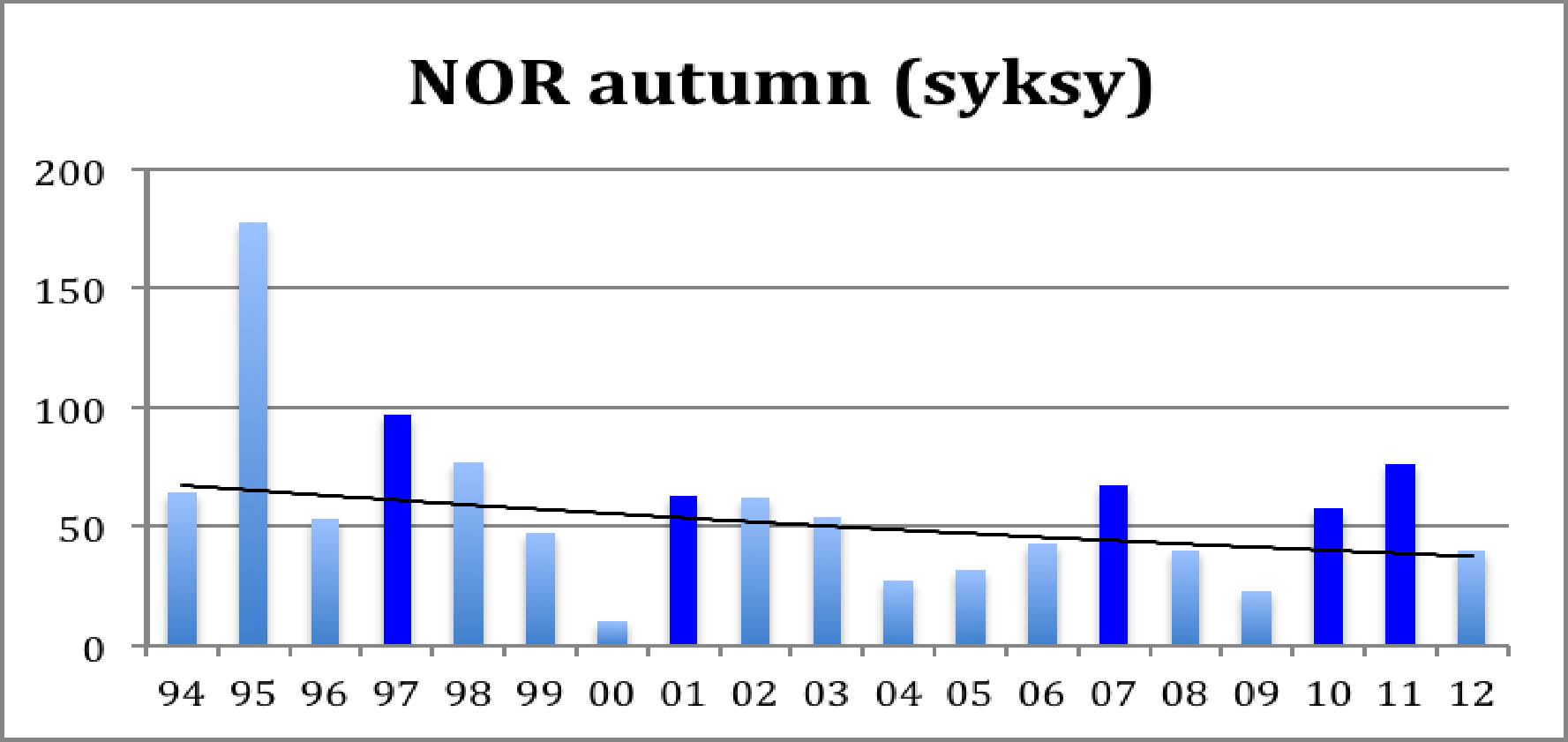
Diagramma 1: Syyslinnut (aikuiset + poikaset)
Ensimmäiseen diagrammaan on merkitty Norjan "Piskulka-sivulta" kootut kiljuhanhien lukumäärät syyskeräytymässä Norjassa. Sopulivuodet on merkitty tummalla. Loivasti laskeva käyrä esittää eksponenttifunktiosovitusta. Prof. Heikki Henttonen Metsäntutkimuslaitokselta on ystävällisesti toimittanut käyttööni tiedot sopulivuosista. Niitä ovat olleet 1937-1938, 1942 (paikallista), 1946 (paikallista), 1959 (paikallista), 1960 (heikkoa), 1969&endash;1970 (aika vahvaa), 1974, 1978, 1982 (heikkoa), 1997-1998 (jäi kesken), 2001 (heikko), 2007 (painopiste Norjassa), ja 2010-2011 (vahvat). Maksimivuosien välit ovat 4, 4, 23-24, 4, 4, 4, 15, 4, 6, ja 3-4 vuotta. Neljän vuoden jaksollisuus on tunnettu asia, mutta pitkät tauot ovat huonommin tunnettuja. Jos nekin ovat nykyisin jaksollisia, on seuraava tulossa, mutta kahden havainnon perusteella on vielä aika uskaliasta (joskin mielenkiintoista) ennustaa mitään.
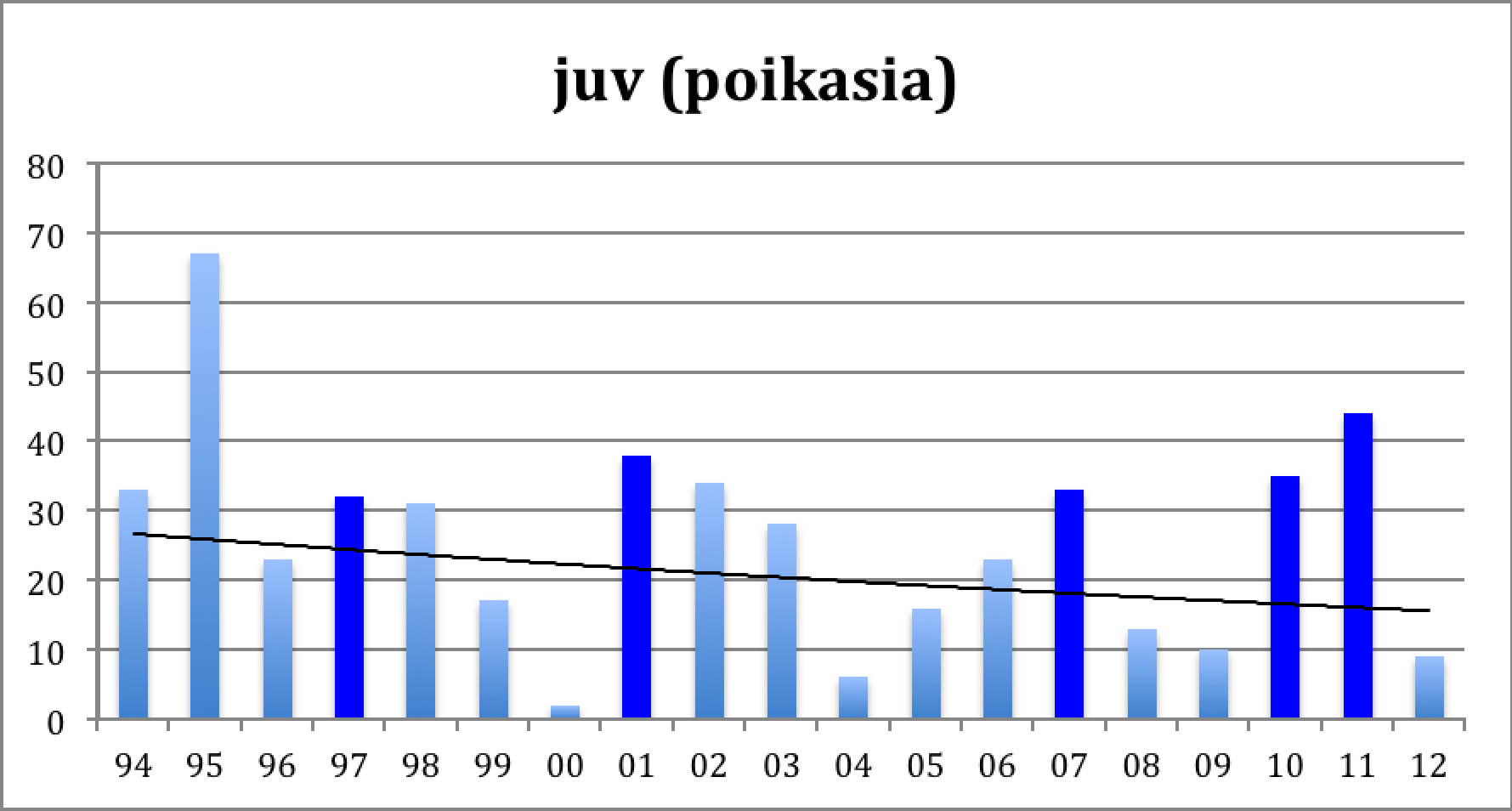
Diagramma 2: Syyspoikaset
Toiseen diagrammaan on merkitty Norjan "Piskulka-sivulta" kootut kiljuhanhien poikasten lukumäärät syyskeräytymässä Norjassa. Sopulivuodet on taas merkitty tummalla ja viiva on eksponenttifunktiosovitus.
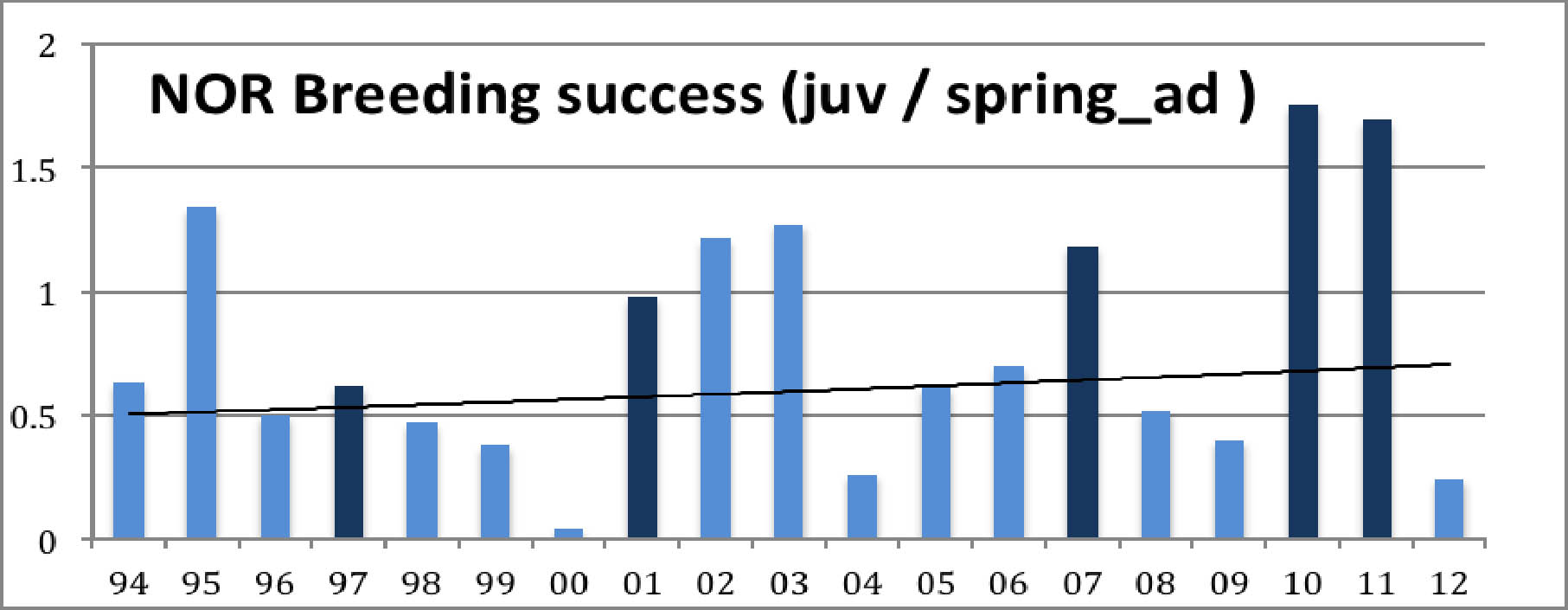
Kummastakin diagrammasta näkyy sama asia, kiljuhanhien väheneminen, jota rytmittävät sopulivuosiin liittyvät tilapäiset kannan toipumiset. Vuoden 2000 pesinnän tuhosi poikkeuksellisen huono sää. Muuten kummastakin kuviosta näkyy hyvä poikastuotto sopulivuosina ja sen päällä loivasti laskeva trendi. Pienellä laskutoimituksella voi todeta, että syyslintujen keskimääräinen väheneminen on noin 3,0 prosenttia vuodessa, mikä johtaa kannan puolittumisen 20 vuodessa. Poikastuoton aleneminen on hieman pienempi, 2,7 % vuodessa. Ero merkitsee, että pesinnät ovat alkaneet onnistua hieman paremmin. Näin asia onkin: Viimeisen vuosikymmenen aikan keskimääräinen pesimätulos on ollut 0,77 poikasta/kevätlintu ja edellisellä vuosikymmenellä vain 0,68. (Koko ajalta 0.71.) Pesimätulosta kannattaa katsoa vuosittain:
Diagramma 3: Pesimätulos poikasia/kevätaikuinen
Poikkeusvuoden 2000 lisäksi huomiota herättää suuri, kasvava ero sopulivuosien ja muiden välillä. Sopulivuosien pesimätulos näyttää paranevan ja muiden vuosien pesimätulos huononevan. Lisätään kaavioon trendiviivat kummallekin:
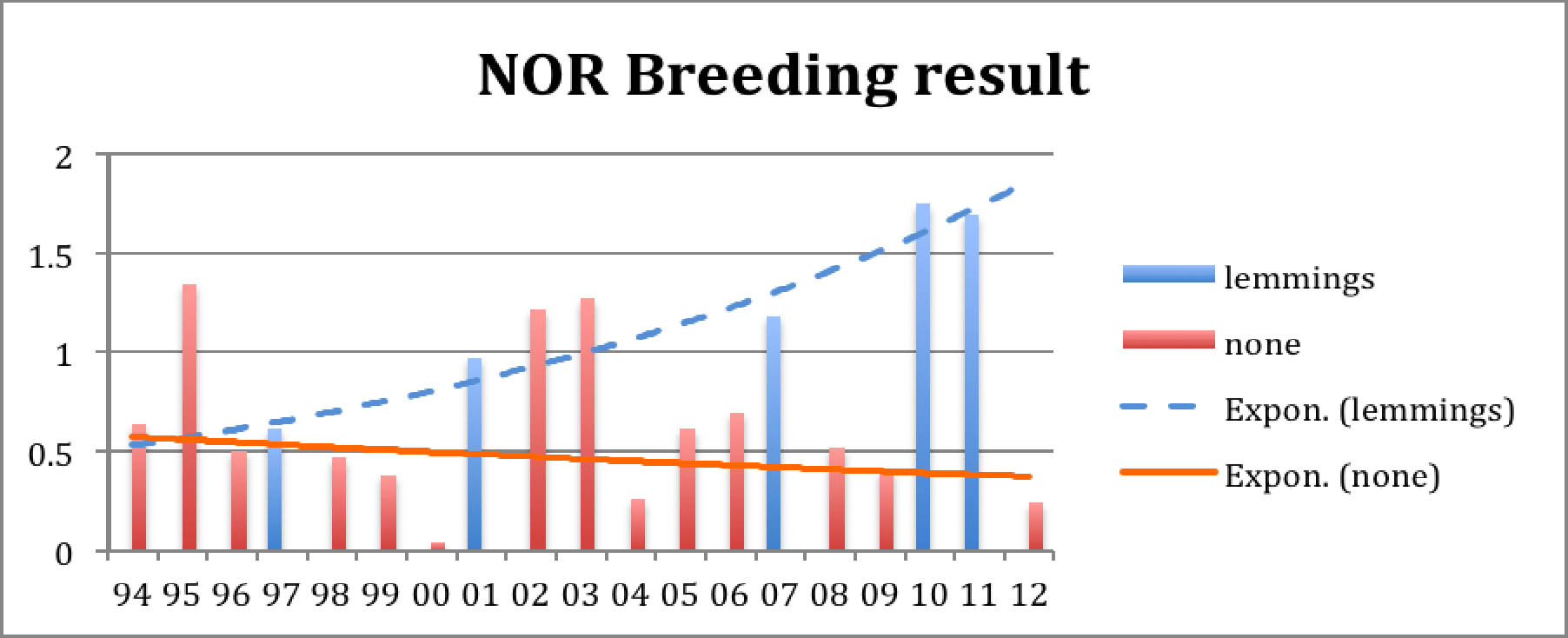
Diagramma 4: Kiljuhanhen pesimätulos sopulivuosina (lemmings) ja muina vuosina
Sopulivuosien yhteys kiljuhanhiin on tunnetusti se, että pienpedot jättävät kiljuhanhet pesimään rauhassa, kun niille riittää syötäväksi helposti saalistettavia sopuleita. Sopulivuosina kiljuhanhien poikastuotto siis on suuri. Tämä näkyy hyvin kummastakin diagrammasta. Voimme nyt tarkastella neljän eri sopulivuoden jälkeistä kiljuhanhikannan romahdusvauhtia ja huomata, että kanta on aina suunnilleen puolittunut huippuvuodesta kahden tai kolmen vuoden jälkeen. Koska kiljuhanhien poikaskuolleisuus on muutolla paljon suurempi kuin aikuisten lintujen ja koska edellisvuoden poikaset ja muutkin pesimättömät tai pesinnässään epäonnistuneet linnut muuttavat itäisempää, pitempää ja vaarallisempaa reittiä kuin syyspoikueet, on tapahtumien kulku helppo arvata: Sopulivuoden syksyllä kiljuhanhien määrä on kaksinkertaistunut keväisestä: puolet syyslinnuista on nyt poikasia, puolet niiden vanhempia. Pesimättömiä aikuisia on vain muutamia. Linnut muuttavat lyhyttä reittiä. Seuraavana keväänä muutolta palaa suurin osa vanhoista emolinnuista ja noin puolet poikasista. Sopulivuoden jälkeisen kesän pesimätulos on huono, koska nälkäisiä pikkupetoja on nyt paljon. Syysparvi muodostuu nyt muutaman poikueen lisäksi pääosin pesimättömistä aikuisista linnuista ja vielä hengissä olevista edellisvuoden poikasista. Kannan koko on siis noin 75 % edellisvuoden huipusta. Nytkin poikueet lentävät edullisinta reittiä, mutta niitä on vähän ja valtaosa kiljuhanhista lentää nyt itään, jossa etenkin edellisvuoden poikaset kuolevat. Toisena keväänä sopulivuoden jälkeen lähes kaikki sopulivuonna kuoriutuneet poikaset ovat kuolleet ja vain eloonjääneet alkuperäiset vanhat linnut palaavat Norjaan mukanaan muutama hyvän vuoden nuori ja muutama huonon vuoden poikanen. Seuraavan kesän pesimätulos on jälleen vain tavanomainen ja kahden vuoden kuluttua huippusyksystä on kanta suunnilleen samankokoinen kuin ennen sopulivuotta. Jos uutta sopulivuolta ei tule, kanta alenee hieman ennen seuraavaa kierrosta, joka toistaa saman kehityksen pienemmällä lintumäärällä. Näin sekä teoriassa että havainnoissa!
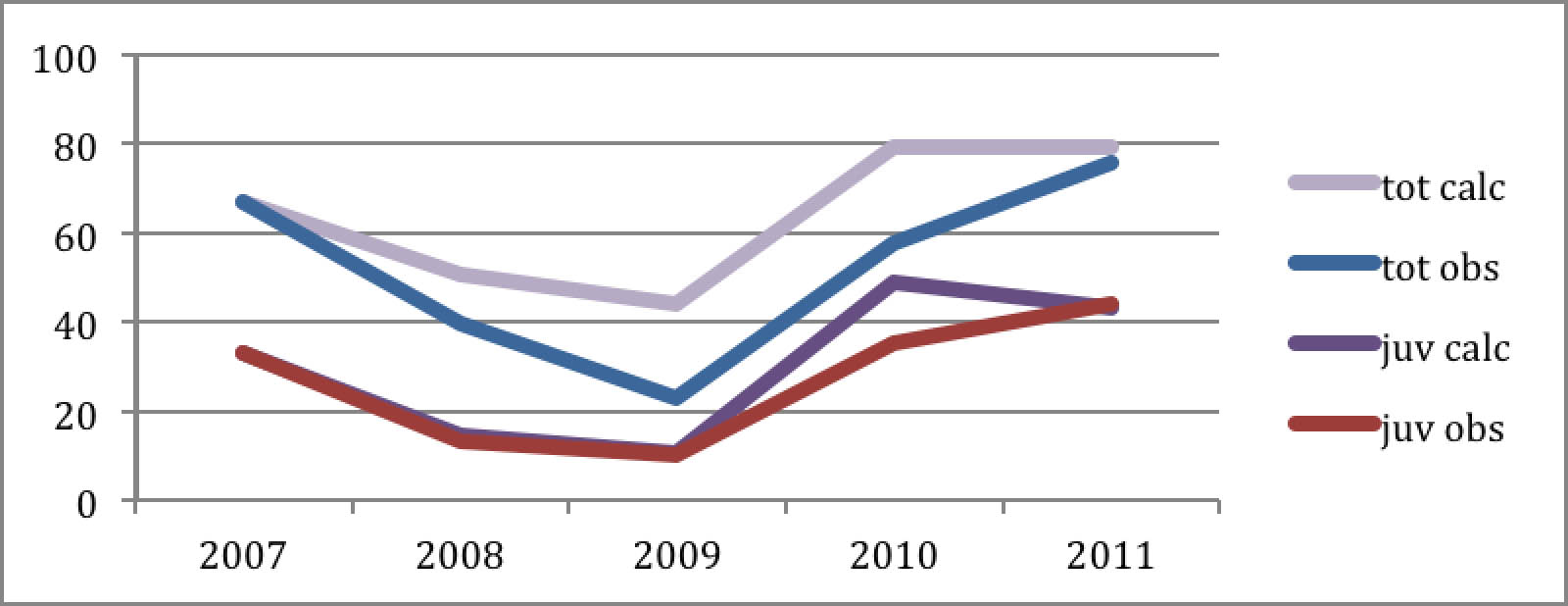
Lasketanpa tarkasti ja yksinkertaisesti: Käytetään laskun pohjana WWF:n (ja minun) laskemia kuolleisuuslukuja 1990-luvulta: nuorten kiljuhanhien ensimmäisen muuton kuolleisuus on sen mukaan 78 % ja aikuisten 16%. Yksinkertaisuuden vuoksi kesäkuolleisuus on huomioitu muuttokuolleisuudessa, joten siitä ei tarvitse välittää. Lisääntymisessä otetaan nyt huomioon sopulivuodet ja lasketaan yhden nelivuotisjakson &endash; vaikkapa viimeisimmän &endash; tulos. Vuoden 2007 syksyllä Norjassa havaittiin 29 aikuista, 5 edellisvuotista ja 33 saman vuoden poikasta. Kuolleisuuden mukaan keväällä 2008 olisi teoriassa jäljellä 24 aikuista, 4 esiaikuista and 7 nuorta. Koska 2008 oli sopulivuoden jälkeinen, pesimätulos oli vain 0,52 poikasta/(esi)aikuinen. Syysparven kooksi tulee näin laskien 51 lintua, nimittäin 29 aikuista, 7 esiaikuista ja 15 poikasta. Laskutoimitusta voidaan jatkaa samalla tavalla seuraaville vuosille. Sopulittomina vuosina 2008 ja 2009 pesimätulosluku oli 0.52 ja 0.36, seuraavina sopulivuosina 2010 ja 2011 taas 1.75 ja 1.69. Käyttämällä näitä lukuja ja samoja vanhoja kuolleisuuslukuja saamme syysparven kooksi seuraavaa:
Diagramma 5: Havaitut (obs) ja lasketut (calc) kaikki (tot) ja nuoret( juv).
Laskun tulos täsmää lähes täydellisesti yhteen havaintojen kanssa &endash; etenkin lopputulos koko 4 vuoden syklin jälkeen. Koska lisääntymistä kuvaavat kertoimet olivat laskelmassa samat kuin havaitut todelliset, niin käyrien yhtenevyys merkitsee, että laskussa käytetyt kuollleisuuskertoimet ovat vaikuttaneet saman kuin todellsetkin. Toisin sanoen vanhat kuolleisuuskertoimet ovat edelleen voimassa &endash; kuolleisuus ei siis ole alentunut edellisestä vuosikymmenestä, vaan muutokset ovat tapahtuneet pesimäpuolella. Viralliset suojelutoimet ovat kuitenkin kohdistuneet melkein yksinomaan talvehtimiseen. Niillä ei siis ole ollut mitään vaikutusta. Pesimätulokseen taas on vaikuttanut ilmeinen tekijä: sopulit.
Tätä kirjoittaessani on loppukevät 2013 ja talvehtivat kiljuhanhet on juuri saatu laskettua niin Kreikassa kuin Unkarissakin. Suurimmat parvet ovat kummassakin paikassa tällä kertaa pienentyneet hieman edellisen periaatteen mukaan odotettua nopeammin: Kreikassa suurimassa parvessa oli edellistalvena 70 kiljuhanhea ja kuluneena talvena 48, Unkarissa vastaavasti noin 70-80 ja 42.
Palaan ennusteisiini ja myönnän heti, etten niitä laatiessani ollenkaan ottanut huomioon sopulivuosia. Ensimmäinen, Viron Roostassa vuoden 2004 hanhikonferenssissa esittämäni ennuste perustui julkaistuihin havaintoihin, joista laskin nuorille ja vanhoille linnuille kuolleisuuden muuttomatkalla ja keskimääräisen pesimätuloksen. Yksinkertainen malli antoi ennusteen, jonka päätulos oli noin viiden prosentin väheneminen vuosittain. Kolme vuotta myöhemmin Odessan konferenssissa saatoin todeta ennusteen toteutuneen hyvinkin tarkasti, mutta ei aivan tasaista vauhtia. Niinpä olin laatinut uuden matemaattisesti aika monimutkaisen ennusteen, joka otti huomioon myös kuolleisuuden satunnaisen vaihtelun. Mallin hienosäädön tulos oli, että vaihtelu lisää sukupuuton riskiä etenkin kannan jo ollessa pieni. Tulos vastasi havaintojen lisäksi myös talonpoikaisjärkeä: suuren kuolleisuuden sattuessa ei pienestä joukosta jää eloon montakaan lintua, ehkei yhtään. Hiljainen hiipuminen vie enemmän aikaa.
Kun vuonna 2007 taas tarkastelin tehtyjä ennusteita, ne pitivät edelleen paikkansa aika hyvin: Norjan kiljuhanhikannan trendi oli 4- 5 % väheneminen. Nyt laadin paljon laajennetun ennusteen, joka ei koske ainoastaan Norjan lintuja vaan myös Ruotsin ja euroopanpuoleisen Venäjän kiljuhanhia sekä tarhalintuja ja Suomeen mahdollisesti istutuksin syntyvää kantaa. Jotta ei uuden havaintoaineiston tullessa tarvitsisi korjailla mallia tein siihen säätöpaikat eri kantojen ja ikäluokkien kuolleisuudelle, pesimätulokselle ja lintujen siirtelylle kannasta toiseen &endash; siis istutusprojekteille. Malli esiteltiin toistaiseksi viimeisessä kansainvälisessä avoimessa tieteellisessä kiljuhanhikonferenssissa Saksan Xanteniissa 2007 ja se julkaistiin vertaisarvioidussa artikkelissa arvostetussa die Vogelwelt-lehdessä. Sitä on sittemmin käytetty myös ulkomaisten tutkijoiden työkaluna. Malli on jokaisen käytettävissä ja oman maun mukaan viritettävissä internet-osoitteessa www.piskulkaconf.tk klikkaamalla " Calculate easily the effects of protection measures on LWfG populations! " ja taustatiedoksi
Background document " . Malli on sen verran laaja, etten ole rakentanut siihen satunnaisen vaihtelun vaikutusta, joka jo Odessassa oli todettu vähäiseksi. Syöttämällä malliin kuolleisuus- ja muut luvut se tuottaa ennusteita. Oletusarvoina syöttämäni pesimätulokset näyttävät uusimpien havaintojen valossa aliarvioiduilta ja siitä syystä näyttää myös kannan hupeneminen antamieni oletusarvojen mukaista ennustetta hitaammalta. Tämä ilmiö selittyy lähes kokonaan sillä, että juuri nyt on takanamme runsaiden sopulien aika. Siksi on liian aikaista sanoa, onko kiljuhanhien vähenemisessä tapahtunut mitään pitkäaikaista muutosta.
Luonnollisesti tarhakannat ja istutuksen vaikutukset sisältävä tietokonemalli kelpaa myös työkaluksi suunniteltaessa kiljuhanhen pelastustoimenpiteitä, etenkin istutusta. Siihen tarkoitukseen malli on tehty ja siihen sitä kannattaa kokeilla. Mutta se on toinen tarina.
Jyrsijäkantojen, etenkin sopulien vaikutus hanhien pesimätulokseen on yleisesti tunnettu asia, mutta merkitessäni sopulivuodet diagrammoihin 1 ja 2 yllätyin kyllä siitä miten voimakas ilmiö on. Kiljuhanhien tulevaisuuden kannalta voi olla hyvinkin olennaista miten tunturisopulien kannanvaihtelut kehittyvät ilmaston-muutoksen myötä. Akuutisti kiljuhanhen kohtalo kuitenkin riippuu vain siitä aloitetaanko sen aktiivinen suojelu eli voimaperäinen tarhaus ja istuttaminen viimeinkin myös Ruotsin ulkopuolella &endash; tähän asti kokeillut muut keinot olen yllä osoittanut tehottomiksi.
Thomas Heinicke ja Lauri Kahanpää (toim.)

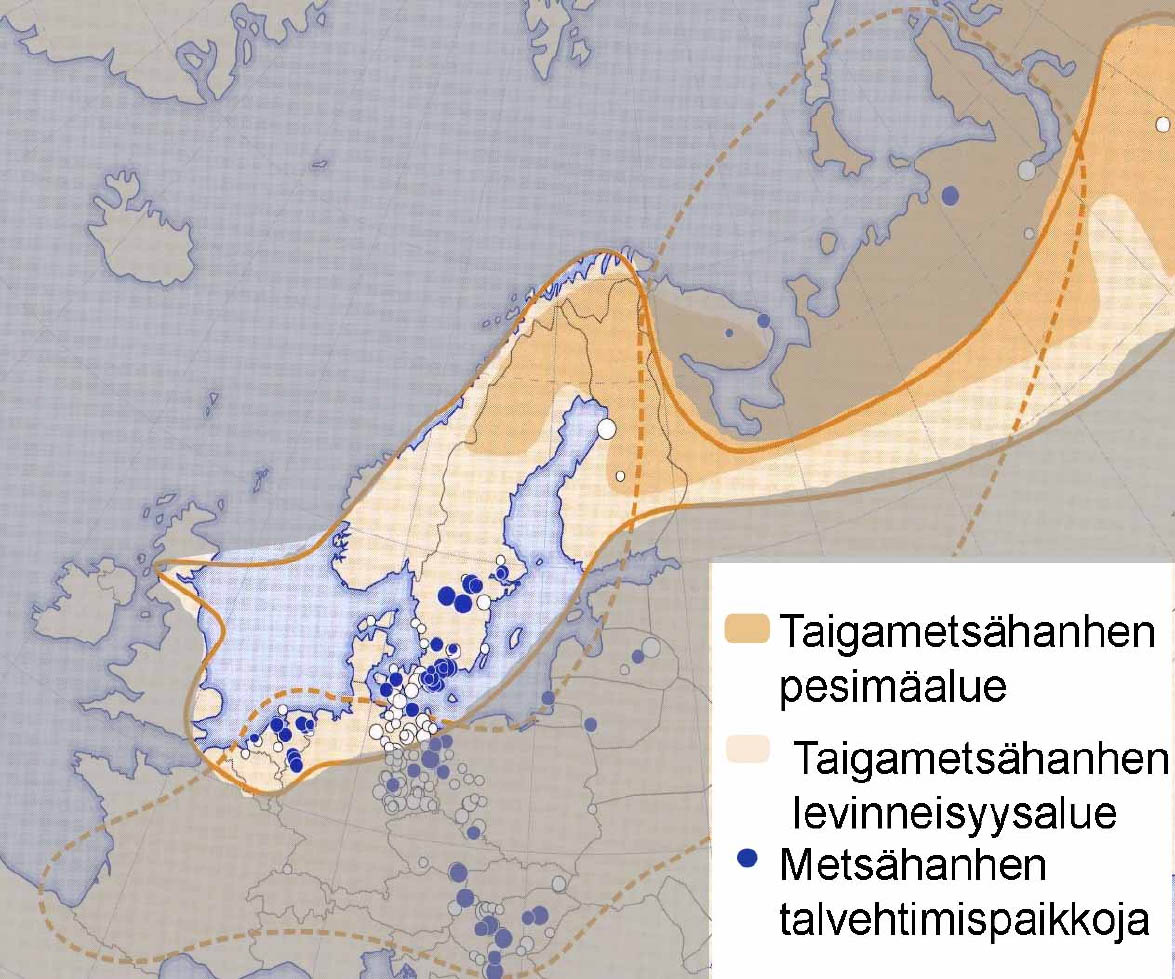
Toisin kuin kiljuhanhella on metsähanhella useita alalajeja, joista nimirotu Anser fabalis fabalis pesii meillä ja alalajia Anser fabalis rossicus tavataan etenkin maamme kaakkoisosassa läpimuuttajana. Englannin kielessä näitä alalajeja on pesimäalueidensa mukaan tapana kutsua taiga- ja tundrametsähanheksi. Tässä kirjoituksessa osoitetaan, että näiden alalajien selvä eriytyminen toisistaan antaa aiheen suojella kumpaakin erikseen, ja että juuri Suomi on avainasemassa uhanalaisen taigametsähanhen suojelussa.
Kirjoitus perustuu Thomas Heinicken GOOSE 2011-konferenssissa pitämään esitelmään ja julkaistaan hänen pyynnöstään kiinnittämään huomiota Suomen metsähanhien erityisasemaan.
Taigametsähanhi on lähes merihanhen kokoinen, sen nokka on pitkä ja hoikka, useimmiten väriltään laajalti oranssi, mutta toisinaan kärkialuetta lukuunotta-matta harhaanjohtavasti tumma, kuten rossicus- alalajilla eli tundrametsähan-hella, joka on pienempi, tummempipäinen ja lyhyempikaulainen. Rossicuksen nokka on tyviosaltaan aina tumma ja paksu. Tarkemmt tunnistusohjeet saa sivulta http://www.birdlife.fi/suojelu/lajit/tundrametsahanhi.pdf .
Metsähanhi on Euraasiassa laajalle levinnyt lintu, mutta nimirotu pesii vain Suomessa, Ruotsissa ja kaistalla Karjalasta Uralille. Metsähanhen ja sen täkäläisten alalajien runsaudesta pääsee selville laskemalla ne talvehtimis-alueillaan. Metsähanhista osa talvehtii tosin piilossa Aasian puolella, mutta useimmat Euroopan metsähanhista, etenkin kaikki Suomessa tavatut, talvehtivat Keski-Euroopassa, jossa ne tosiaan lasketaan.
Kun metsähanhien laskennassa on alettu kiinnittää tarkkaa huomiota alalajeihin, on Euroopan talvehtimisalueilla huomattu, että Suomessa pesivä taigametsähanhi on äkkiä alkanut harvinaistua. Sen talvilaskennoissa on saatu seuraavat tulokset:
Tammikuu
S
DK
D
PL
UK
Yht.hav
Arvio
2004
9.326
10.683
35.000
3.800
375
69.200
70.000-90.000
2005
34.560
8.728
42.000
490
418
86.200
70.000-90.000
2006
19.289
16.279
52.000
1.500
469
89.500
70.000-90.000
2009
32.500
13.836
22.500
1.500
471
70.800
60.000-65.000
2011
8.201
20.000
12.100
1.790
453
42.544
45.000
Kymmenessä vuodessa nimirodun kanta on siis puolittunut ja kahdessa vuodessa (2009-2011) se on menettänyt yli 20 000 lintua! Mistä näin äkillinen romahdus voi johtua? Onko vika pesinnässä vai kuolleisuudessa? Pesinnän onnistumista seurataan yleensä laskemalla nuorten lintujen osuus syysparvissa. Tällainen laskenta tehtiin keski-Ruotsissa syksyllä 2009 ja uudelleen 2010. Laskennassa ei erotettu alalajeja. Tulos on seuraava:
Aika
nuoria
yhteensä
poikuekoko
poikueita
Syyskuu 2009
22,9%
6.710
2,56
209
Lokakuu 2009
20,90%
3.727
Syyskuu 2010
26,1%
4.202
Lokakuu 2010
17,2%
1.538
2,49
160
Marraskuu 2010
7,7%
2.521
Alkusyksystä nuoria lintuja on paljon, jopa hieman enemmän kuin arktisilla hanhilla yleensä. Myöhemmin syksyllä nuorten osuus alenee huomattavasti. Täl-lainen ilmiö voisi johtua siitä, että pesimättömät ja pesinnässään epäonnistuneet linnut muuttaisivat eri aikana tai eri reittiä kuin pesinnässään onnistuneet. Toinen selitys olisi, että syksyn edetessä keski-Ruotsiin tulisi metsähanhia idästä, ja niillä olisi eri pesimätulos tai matkan varrelta (Suomesta päin!) jo nuorten lintujen tappioita toisin kuin paikallisemmilla. (Toimittajan huomautus: Metsästyksen vaikutus Ruotsissa ei ilmeisesti ole syy. Pohjois- ja keski-Ruotsissa metsähanhia ei metsästetä, etelässä kyllä.)
Lisätietoja kuolleisuudesta antavat pohjois-Ruotsissa ja Saksassa tehdyt kaularengasmerkinnät ja havainnot:
Rengastusvuosi
lkm
% elossa 2006/2007
% elossa 2007/2008
% elossa 2008/2009
% elossa 2009/2010
% elossa 2010/2011
Kevät 2007
1
100
100
100
100
100
Kevät 2008
27
100
85,2
77,8
51,9
Kevät 2009
11
100
72,7
45,5
Kuolleisuus Ruotsissa
14,3 (4/28)
14,3(5/35)
33,3(10/30)
Saksa (Oderin alajuoksu) syksy 2007
33
100
57,6
33,3
12,1
Kuolleisuus Saksassa
42,4
42,1
63,6
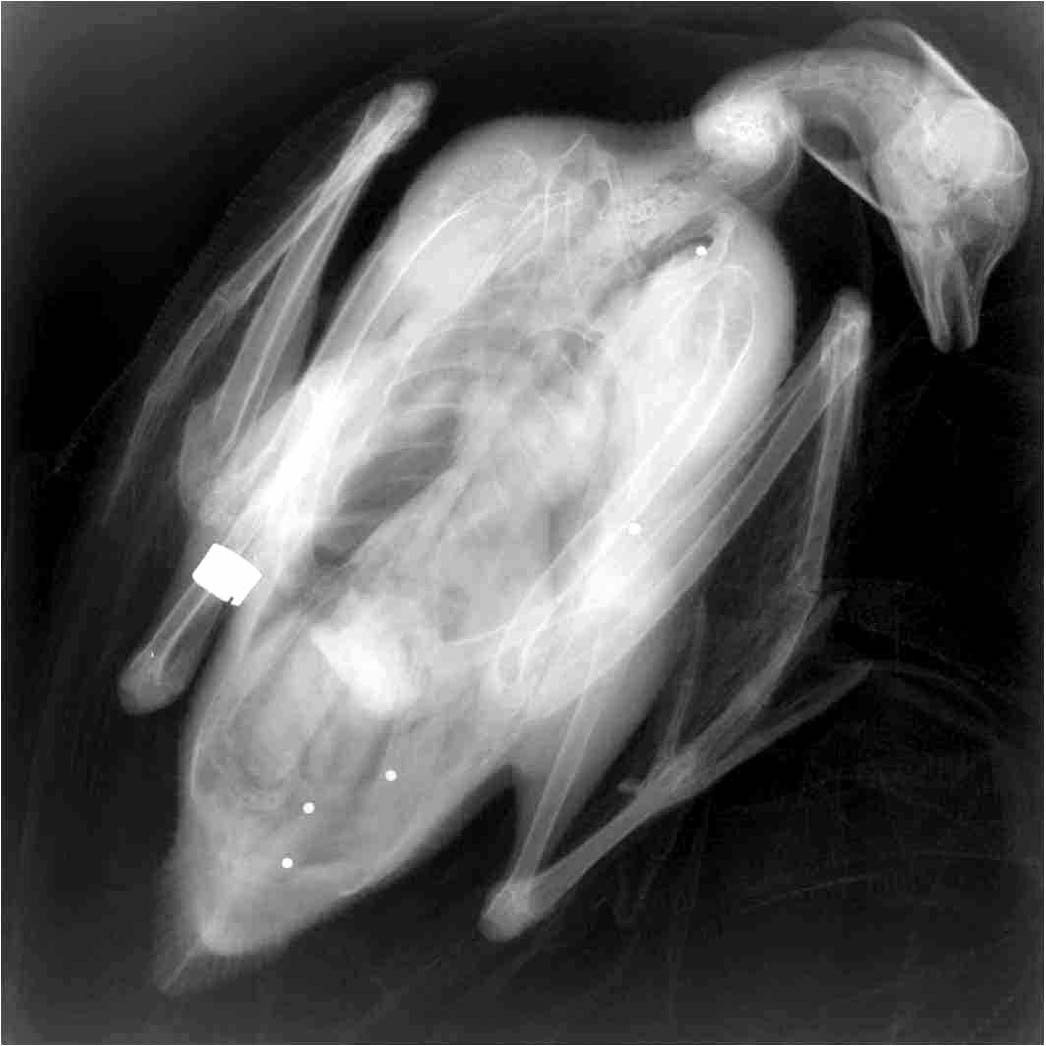
Huomiota herättää, että kaularengastettujen metsähanhien kuolleisuus on itäisessä Saksassa yli kaksinkertainen pohjois-Ruotsiin verraten. Metsästyksen osuutta tähän ilmiöön voi arvioida tutkimalla kuinka monessa linnussa on hauleja. Ne näkyvät röntgenkuvassa: (Kuvassamme 5 kpl.)
Seuraavassa taulukossa näkyy hauleja kantavien hanhien osuus itä-Saksassa (ala)lajeittain ja iän mukaan.
nuoria puhtaita
nuoria haulein
aikiuisia puhtaita
aikuisia haulein
haulein yht
merihanhi
1
0=0%
58
14=19,4%
19,2
tundrahanhi
35
0=0%
73
20=21,5%
15,6
tundra-metsähanhi
30
0=0%
44
24=35,3%
24,5
taiga-metsähanhi
4
0=0%
13
11=45,8%
39,3
21,1
Huomiota herättää, että itäisessaä Saksassa tehdyissä röntgenkuvauksissa nähtiin ylivoimaisesti eniten hauleja aikuisissa taigametsähanhissa eikä minkään lajin nuorissa linnuissa nähty yhtään. Ammutaanko lintuja siis vasta Saksassa? Metsästystilasto (Siteerattu Hirschbach & Heyd 2005) kertoo seuraavaa mestähanhista:
saalis
tilastovuosi
Huom
Norja
0
rauhoitettu
Suomi
7.900
2009
Pääosin taiga-
Ruotsi
3.450
2005/2006
Pääosin taiga-
Tanska
886
2005/2006
Pääosin taiga-
Saksa
4.255
2005/2006
Pääosin tundra 300-500 taiga
Puola
13.812
2005/2006
Pääosin tundra 200-400 taiga
Baltian maat
1.127
2005/2006
Pääosin tundra 100-200 taiga
V-venäjäja Ukraina
?
Pääosin tundra 200-400 taiga
Venäjä
5.000-10.000
Vähintään
Yhteensä
35.000-40.000
Tässä on siis suunnilleen se mitä todella tiedetään taigametsähanhesta:
1. Keski-Euroopassa talvehtimisalueilla hiljattain vähentyneet yhä nopeammin.
2. Tällä menolla häviävät noin 5 vuodessa.
3. Täyttää kirkkaasti IUCN-ehdon (A2) ollakseen maailmanlaajuisesti uhanalainen: viimeisten 10 vuoden (tai 3 sukupolven) aikana se on vähentynyt yli 30 %
4. Ovat todella vaarassa kuolla ilman nopeita toimia.
5. Syyt vähenemiseen: liika metsästys (Suomi! Venäjä? Puola? Saksa?) ja elinympäristöjen väheneminen (Suomi! Venäjä?)).
Kuva: Hämeenkosken kiljuhanhitarhan yhteydessä toimivassa lintuhoitolassa on muutamia metsähanhiakin. Tällä menolla on olemassa vaara, että joudumme palauttamaan istutuksin Suomeen molemmat lajit!
Sukupuuttoon kuolleista lintulajeista:
Lauri Kahanpää
Lintujen luokka Aves on tuottanut uusia lajeja jo noin sadan miljoonan vuoden ajan. Kaikkien yksilöiden kuolema, lajin sukupuutto, on aikojen kuluessa kohdannut niistä useimpia ja parhaillaan on menossa yhtäaikaisten suku-puuttojen aalto: vuoden 1500 jälkeen alkaen on hävinnyt lopullisesti noin 200 lintulajia, siis enemmän yksi lintulaji joka kolmas vuosi. Ainakin 91 lintulajia on kuollut sukupuuttoon vuoden 1681 jälkeen. Internet-sivu Ornithology.com Home sisältää luettelon näistä sekä pahasti vaarantuneista ja varmasti tai todennäköisesti sukupuuttoon kuolleista lajeista. Englanninkielisen nimen lisäksi mainitaan lajin tieteellinen nimi, sukupuuton aika ja paikka ja onko kyseessä laji vai alalaji sekä status.
Euroopassa pesivistä linnuista ei kuitenkaan ole tuona aikana kokonaan hävinnyt ainoakaan laji. On Suomenkin vastuulla huolehtia siitä, ettei kiljuhanhesta tule ensimmäinen.
Solnhofenin liuskeen Archeaopteryx oli kauan lähes ainoa tunnettu vanha lintufossiili. Dinosaurusten aika, mesotsooinen maailmankausi, muodostuu trias- jura- ja liitukaudesta ja päättyi noin 60 miljoonaa vuotta sitten. Solnhofenin liuske on peräisin jo jurakau-delta, siis peräti 200&endash;150 miljoonaa vuotta vanhaa. Hiljattain tehdyt uudet fossiililöydöt ovat varmistaneet lintujen luokan korkean iän. Yksi näistä löydöistä on oheisen kuvan esittämä Confuciusornis, jolla toisin kuin samanikäisellä Archeaopteryxillä on hampaaton nokka. Linnut ovat näin ollen paitsi varhaisten dinosaurusten jälkeläisiä myös myöhempien dinosaurusten ja lento-liskojen aikalaisia. Liitukaudelta tunnetaankin useita lintulahkoja, kuten Enantiornithes, Hesperornithiformes, ja Ichthyornithiformes ja tiedetään, että liitukauden lintuharrastaja olisi lento- ja hirmuliskojen seassa hyvinkin voinut havaita myös nykylintujen (Neornithes) esi-vanhempia, joista erottui jo mm. lokkeja, kahlaajia ja sorsalintujakin. Toisin kuin muut dinosaurukset ja linnut, nykylinnut selvisivät hengissä kenotsooisen maailman-kauden aloittaneesta massatuhosta ja tertiäärikauden aikana eriytyivät nykyiset lintuheimot. Jo oligoseenikaudella noin 30 miljoona vuotta sitten oli olemassa niistä nuorinkin, menestyksekkäät varpuslinnut. Tertiääriajalle kuuluvat myös ne nykyisten strutsien kokoiset lentokyvyttömät petolinnut, vai pitäisikö sanoa lintupedot, joita kehittyi toisistaan riippumatta eri maanosissa. Kvartäärin, siis jää- ja lämpökausien vaihtelun alkaessa pari miljoonaa vuotta sitten ne olivat jo kauan olleet hävinneinä, mutta maailman linnusto ei kuitenkaan ollut täysin nykyisen kaltainen. Kehitykseen olivat vaikuttaneet monet luonnolliset syyt, kuten ilmaston muutokset ja muiden lajien ja lajiryhmien aiheuttama kilpailupaine. Esimerkiksi tuona aikana tapahtuneen merilintujen monimuotoisuuden romahduksen on ilmeisesti aiheuttanut merinisäkkäiden runsastuminen. Sukupuuttoaaltojen syitä ovat myös luonnonkatastrofit kuten suuret tulivuorenpurkaukset ja meteori-iskut.
Kaikki nämä ilmiöt jatkuvat tietenkin edelleen eivätkä jääkaudet ainakaan ole olleet omiaan vähentämään sukupuuttoja. Viimeisen miljoonan vuoden aikana sukupuuttoon kuolleita fossiilisia lintulajeja on nimetty pitkälti yli 300 ja paljon on tuntemattomiakin, sillä linnut ovat luustoltaan hauraita, fossiloituen huonosti tai vain osittain eikä kokonaisia luurankoja löydetä usein muista jäännöksistä puhumattakaan.
Vasta viimeisen jääkauden aikaan osuu kiljuhanhen eriytyminen tundrahanhesta samoin kuin monen muunkin lajiparin muodostuminen. Nykyisen lämpökauden aikana on havaittavissa ihmisen vaikutus luontoon, myös lintujen sukupuuttoihin. Yleisesti tiedetään, että esimerkiksi Uuden Seelannin lentokyvyttömät jättilinnut moat tapettiin sukupuuttoon jo maorien toimesta, mutta varhaisen ihmisen muu vaikutus lintuihin on silti edelleen huonosti tunnettua. Hyvin tunnettua on sen sijaan linnuston kehitys viimeisen 500 vuoden ajalta, jolta on olemassa tieteellisiä muistiinpanoja ja yritystä lajiston selvittämiseksi. Tulos on häkellyttävä: Vuoden 1500 jälkeen alkaen on hävinnyt lopullisesti noin 200 lintulajia, siis enemmän yksi lintulaji joka kolmas vuosi. Tämä merkitsee, että meneillään on nopea massa-sukupuuttojen aika. Syy on varmasti ihmisen, joka aiheuttaa sukupuuttoilmiöitä monin tavoin. Kiljuhanhen liiasta metsästyksestä tietoisina meille tulee varmaan ensin mieleen juuri kestämätön metsästys tai muu tappaminen.
Ruokapöydän kautta ovat hävinneet niin siivetön ruokki kuin dodo eli dronttikin, mutta myös muuttokyyhky, joka ei kestänyt parviensa pienentämistä alle miljooniin yksilöihin. Pienen parvien suojattomuus saattaa vaivata kiljuhanheakin. Muista ihmisen aiheuttamista ongelmista ajankohtainen on ilmastonmuutos, jonka vaikutus lintuihin on sekin hyvin havaittava &endash; itse asiassa juuri lintujen levinneisyyden muutokset ovat hyvin selvästi havaittava ja herkkä indikaattori ilmaston vaihteluille, koska ne liikkuvat niin helposti ja ovat niin näkyviä. Tämäkin koskee kiljuhanhea, kuten eri jutussa tässä lehdessä kerrotaan. Useimmat ihmisen aiheuttamat lintusukupuutot ovat kuitenkin syntyneet muusta syystä kuin välittö-mästä tappamisesta tai elinympäristön tuhoutumisesta, nimittäin haitallisten vieraslajien levittämisestä. Klassinen esimerkki on tarina majakanvartijasta, joka eräänä aamuna löysi eristyneellä pienellä saarella olevan majakkansa rappusilta kuolleen linnun, joka edusti tieteelle tuntematonta lajia. Vartijan kissa oli löytänyt uuden lentokyvyttömän lintulajin ja tappoi sen sukupuuttoon. En ole tarkastanut lajia enkä edes tarinan todenperäisyyttä, mutta se kuvaa joka tapauksessa todellista ilmiötä, sukupuuttojen kohdistumista ennen kaikkea eristettyihin alueisiin, etenkin saariin. Itse asiassa noin 30 % historiallisena aikana kuolleista lintulajeista pesi alun perin Havaijin saarilla ja toisaalta 10 Guamin saaren 13 kotoperäisestä lintulajista on kuollut kolmessakymmenessä vuodessa sukupuuttoon saarelle tuodun ruskopuukäärmeen hävittäminä. Vieraslaji on määritelmällisesti ihmisen levittämä laji, joka ei luonnostaan esiinny jollain alueella eikä voisi sinne levitäkään, koska välissä on maantieteellinen este, kuten valtameri tai ylittämätön vuoristo. Esimerkiksi Suezin kanavan avaaminen poisti tällaisen esteen ja aiheutti monien vieraslajien leviämisen kumpaankin suuntaan. Toinen esimerkki on valkoposkihanhi, joka ei siis ole ihmisen levittämänäkään vieraslaji Suomessa, koska se muuttaa säännöllisesti maamme yli ilman mitään esteitä.
Wikipediassa sivulla http://fi.wikipedia.org/wiki/Luokka:Sukupuuttoon_kuolleet_linnut on suomenkielinen luettelo viimeisen 500 vuoden aikana hävinneistä lintulajeista. Yksikään näistä lajeista ei ole eurooppalainen. Syyksi voi arvata, että Eurooppa on niin kauan ollut niin tiheään asuttu, että todellinen tuho on jäänyt rekisteröimättä, koska se ehti tapahtua ennen varsinaisen lintutieteen alkua jo silloin, kun maatalous levisi ja suuret lehtimetsät muutettiin pelloiksi ja niityiksi. Uusia sukupuuttoja voi kuitenkin tulla pian Eurooppaankin. Koko maailman noin 10 000 lintulajista suunnilleen 12% katsotaan sukupuuton uhan alaisiksi. Sama suhdeluku pätee Euroopassakin, maanosamme 524 lintulajista 68 eli 13% on luokiteltu uhanalaiseksi. Kriittisimmin vaarantunut on kaitanokkakuovi ja listassa on tietenkin mukana myös kiljuhanhi, jonka on Euroopan Unioni on 2011 luokitellut kategoriaan VU asteikolla
- ¥ Arvioimatta jätetyt (NE)
- ¥ Puutteellisesti tunnetut (DD)
- ¥ Hävinneet (RE)
- ¥ Luonnosta hävinneet (EW)
- ¥ Äärimmäisen uhanalaiset (CR)
- ¥ Erittäin uhanalaiset (EN)
- ¥ Vaarantuneet (VU)
- ¥ Silmälläpidettävät (NT)
- ¥ Elinvoimaiset (LC, Least Concern).
Jos nykyinen kehitys jatkuu, on lyhyen ajan kysymys ennen kuin kiljuhanhi on luokassa RE (merkintä tarkoittaa oikeastaan Regionally extinguished, siis alueellisesti sukupuutossa), luokassa EW (vain eläintarhoissa, siis) tai, jos tarhausta ei vieläkään aleta tukea voimakkaasti, kokonaan sukupuutossa &endash; mahdollisesti ensimmäisenä Euroopan lintulajeista historiallisena aikana. Kiljuhanhen pelastaminen tältä kohtalolta on Kiljuhanhen ystävät ry:n päämäärä.
- Postiosoite: c/o Kahanpää / Sepeteuksentie 19 B / FIN 40760 Helsinki
- Kotisivu: http://www.ansererythropus.tk/
- Puheenjohtaja fil.tri. Antti Haapanen
- puh: 358-9-3253804 (k), 358-40-5953313 (m)
- Huhtasuontie 7, 00950 Helsinki, Finland
- e-posti: antti.haapanen@kolumbus.fi
- Sihteeri fil.tri. Lauri Kahanpää
- e-posti: lauri.kahanpaa@jyu.fi
- puh:(0)50-4652301
- Yhdistyksen jäsenyyttä voi hakea sähköpostitse.
- henkilöjäsenmaksu (Suomi) 50 EUR
- yhteisöjäsenmaksu 90 EUR
- kannatusjäsenmaksu 200 EUR
- Pankkiyhteys: S-pankki, 393900-32848780